Toto jsou hlavní hemisféry. Funkce mozkové hemisféry předního mozku
Velké polokoule velké polokoule
mozek, párové útvary spojené corpus callosum do tzv. telencephalon. Povrch mozkové hemisféry reprezentované četnými velkými nebo malými hlubokými konvolucemi. Existují laloky: čelní, parietální, temporální, inzulární, okcipitální. Šedá hmota mozková, tvořená nervovými buňkami – neurony, tvoří mozkovou kůru a podkorové ganglia (uzliny). Bílá hmota je tvořena procesy neuronů, které tvoří mozkové dráhy.
VELKÉ POLOSFÉRYVELKÉ POLOSFÉRY mozku, párové útvary spojené corpus callosum (cm. Corpus Callosum) v tzv telencephalon. Povrch mozkových hemisfér je reprezentován četnými velkými či malými hlubokými konvolucemi. Existují laloky: čelní, parietální, temporální, inzulární, okcipitální. Šedá hmota mozková, skládající se z nervových buněk - neuronů, tvoří mozkovou kůru a podkorové ganglia (cm. GANGLION)(uzly). Bílá hmota je tvořena procesy neuronů, které tvoří mozkové dráhy.
encyklopedický slovník . 2009 .
Podívejte se, co jsou „velké hemisféry“ v jiných slovnících:
Mozek má párové útvary, spojené corpus callosum v tzv. telencephalon. Povrch mozkových hemisfér je reprezentován četnými velkými či malými hlubokými konvolucemi. Existují laloky: čelní, parietální, temporální, ostrovní... Velký encyklopedický slovník
Mozek, párové útvary spojené corpus callosum v tzv. telencephalon. Povrch B. p. je zastoupen četnými. b. nebo m. hluboké konvoluce. Existují laloky: čelní, parietální, temporální, inzulární, okcipitální. Šedá v...... Přírodní věda. encyklopedický slovník
VELKÉ HEMISféry MOZKU- vyšší části mozku, sestávající z povrchové vrstvy mozkové kůry a hlubokých částí subkortexu; pokrývat mozeček a mozkový kmen. B. p. g. m. se dělí podél střední čáry na pravou a levá hemisféra, které jsou v hlubinách... Psychomotorika: slovník-příručka
Severní ledový oceán má na rozdíl od jižního oceánu zcela středomořskou povahu. Má přirozené hranice na značnou vzdálenost a pouze na třech místech přímo splývá s vodami Atlantiku a Pacifiku... ...
Severní ledový oceán má na rozdíl od toho jižního zcela středomořskou povahu. Má přirozené hranice na značnou vzdálenost a pouze na třech místech přímo splývá s vodami Atlantiku a Pacifiku... ... Encyklopedický slovník F.A. Brockhaus a I.A. Efron
Také se nazývá srovnávací morfologie a jedná se o studium vzorců struktury a vývoje orgánů pomocí srovnávání různé typyŽíjící bytosti. Data srovnávací anatomie tradiční základ biologické klasifikace. Podle morfologie... Collierova encyklopedie
Pohled v řezu na mozek dospělého muže. Lidský mozek (lat. encephalon) je o ... Wikipedii
Věda, která studuje stavbu těla jednotlivé orgány, tkání a jejich vztahů v těle. Všechny živé věci se vyznačují čtyřmi vlastnostmi: růst, metabolismus, podrážděnost a schopnost reprodukce. Kombinace těchto vlastností...... Collierova encyklopedie
Živočichové (Mammalia), třída obratlovců, nejvíce slavná skupina zvířat, včetně více než 4 600 druhů světové fauny. Patří sem kočky, psi, krávy, sloni, myši, velryby, lidé atd. V průběhu evoluce si savci uvědomili nejširší... ... Collierova encyklopedie
I Medicína Medicínský systém vědecké znalosti a praktické činnosti, jejichž cílem je upevnění a zachování zdraví, prodloužení života lidí, prevence a léčba lidských nemocí. Aby M. splnil tyto úkoly, studuje strukturu a... ... Lékařská encyklopedie
Mozkové hemisféry jsou největší oblastí mozku. Lidské hemisféry velký mozek dostalo maximálního rozvoje ve srovnání s ostatními částmi, což výrazně odlišuje lidský a zvířecí mozek. Levá a pravá hemisféra mozku jsou od sebe odděleny podélnou trhlinou probíhající podél střední čáry. Pokud se podíváte na povrch mozku shora a ze strany, můžete vidět štěrbinovitou prohlubeň, která začíná 1 cm za středem mezi předním a zadním pólem mozku a jde hluboko. Jedná se o centrální (rolandský) sulcus. Pod ním, podél laterálního povrchu mozku, probíhá druhá velká štěrbinová (Sylviova) štěrbina. Funkce Cerebrální hemisféra přední mozek- téma článku.
1 106654
Fotogalerie: Funkce mozkové hemisféry předního mozku
Mozkové laloky
Velké hemisféry jsou rozděleny na laloky, jejichž názvy jsou dány kostmi, které je kryjí: Čelní laloky se nacházejí před Rolandovou trhlinou a nad Sylviovou trhlinou.
Parietální lalok leží za centrálním lalokem a nad zadní částí laterální sulcus; sahá zpět k parietookcipitálnímu žlábku - oddělující mezeru parietální lalok z týlního hrbolu, který tvoří zadní část mozku.
Spánkový lalok je oblast nacházející se pod Sylviovou trhlinou a ohraničená zezadu týlní lalok y
Jak mozek rychle roste ještě před narozením, mozková kůra začíná zvětšovat svůj povrch a tvoří záhyby, což vede k vytvoření charakteristického vzhled mozek připomínající Vlašský ořech. Tyto záhyby jsou známé jako gyri a prohlubně, které je oddělují, se nazývají trhliny. Určité rýhy se nacházejí u všech lidí na stejném místě, takže se používají jako orientační body pro rozdělení mozku na čtyři laloky.
Vývoj gyri a sulci
Brázdy a konvoluce se začínají objevovat ve 3.–4. měsíci vývoje plodu. Do té doby zůstává povrch mozku hladký, jako mozek ptáků nebo obojživelníků. Vytvoření složené struktury poskytuje zvětšení povrchové plochy mozkové kůry v podmínkách omezeného objemu lebky. Různé oblasti mozkové kůry plní specifické, vysoce specializované funkce. Mozkovou kůru lze rozdělit do následujících oblastí:
Motorické oblasti – iniciují a řídí pohyby těla. Primární motorická oblast řídí dobrovolné pohyby opačné strany těla. Přímo před motorickou kůrou je tzv. premotorická kůra a třetí oblast, doplňková motorická, leží na vnitřní ploše čelního laloku.
Smyslové oblasti mozkové kůry vnímají a syntetizují informace ze smyslových receptorů v celém těle. Primární somatosenzorická oblast přijímá informace z opačné strany těla ve formě impulsů ze smyslových receptorů pro hmat, bolest, teplotu a polohu kloubů a svalů (proprioceptivní receptory).
Povrch lidského těla má svá „zastoupení“ ve smyslových a motorických oblastech mozkové kůry, které jsou určitým způsobem organizovány. Kanadský neurochirurg Wilder Penfield, který praktikoval v 50. letech, vytvořil druh mapy smyslové zóny mozkové kůry, které vnímají informace z různých částí těla. V rámci svého výzkumu prováděl experimenty, při kterých žádal člověka v lokální anestezii, aby popsal své pocity v okamžiku, kdy stimuluje určité oblasti povrchu mozku. Penfield zjistil, že stimulace postcentrálního gyru vyvolala hmatové vjemy ve specifických oblastech na opačné straně těla. Jiné studie ukázaly, že objem motorické kůry, která je zodpovědná za různé oblasti lidského těla, je ve větší míře závisí spíše na úrovni složitosti a přesnosti prováděných pohybů než na síle a objemu svalová hmota. Mozková kůra se skládá ze dvou hlavních vrstev: šedá hmota - tenká vrstva nervových a gliových buněk o tloušťce asi 2-A mm a bílá hmota, který je tvořen nervovými vlákny (axony) a gliovými buňkami.
Povrch mozkových hemisfér je pokryt vrstvou šedé hmoty, jejíž tloušťka se v různých částech mozku pohybuje od 2 do 4 mm. Šedá hmota je tvořena těly nervových buněk (neuronů) a gliových buněk, které plní podpůrnou funkci. Ve většině mozkové kůry pod mikroskopem šest oddělené vrstvy buňky.
Neurony mozkové kůry
- Pyramidové buňky dostaly svůj název podle tvaru těla neuronu, které připomíná pyramidu; jejich axony (nervová vlákna) vystupují z mozkové kůry a přenášejí informace do jiných částí mozku.
- Nepyramidové buňky (všechny ostatní) jsou určeny k vnímání a zpracování informací z jiných zdrojů.
Tloušťka šesti vrstev buněk, které tvoří mozkovou kůru, se velmi liší v závislosti na oblasti mozku. Německý neurolog Korbinian Brodmann (1868-191) zkoumal tyto rozdíly barvením nervových buněk a jejich prohlížením pod mikroskopem. Výsledkem Brodmannova vědeckého výzkumu bylo rozdělení mozkové kůry na 50 samostatných oblastí na základě určitých anatomických kritérií. Následné studie ukázaly, že takto identifikovaná „Brodmannova pole“ hrají specifickou fyziologickou roli a mají jedinečné způsoby interakce.
Mozek (pokračování)
Největší velikost a složitost u savců je přední mozek neboli telencephalon, sestávající ze dvou mozkových hemisfér (hemispheri cerebri). Hemisféry zřejmě vznikly především (a možná výhradně) v souvislosti s čichovým příjmem. Pachy v životě vyšších primátů, včetně člověka, mnoho neznamenají. Avšak v dřívějších fázích evoluce, až po předky obratlovců, byl čich hlavním kanálem, jehož prostřednictvím zvířata získávala informace o světě kolem sebe. Proto je zcela přirozené, že čichová centra mozku sloužila jako základ, na kterém se následně vyvíjely složité nervové mechanismy. Již v raných fázích evoluce tetrapodů se hemisféry mění na velká a důležitá centra pro korelaci smyslových signálů. V době, kdy se objevili savci, se značně rozšířený povrch hemisfér stal dominantním asociačním centrem, místem vyšší mentální aktivity. U různých zástupců třídy se poměr hmotnosti hemisfér předního mozku k hmotnosti celého mozku liší: u ježka ( Erinaceus europaeus) je to 48 %, u veverek ( Sciurus vulgaris) - 53 %, u vlka ( Canis lupus) - 70 %, v bílé barvě ( Delphinus delphis) - 75 %, u většiny primátů - 75-80 %, u lidí - asi 85 %. U ptáků mají mozkové hemisféry přibližně stejnou hmotnost jako zbytek mozku nebo jsou pod ním, někdy i několikrát. Konečně o mimořádném významu mozkových hemisfér svědčí fakt, že jejich destrukce vede k úplnému funkčnímu selhání savce.
Čichové bulby (bulbi olfactorii) přiléhají k přední části hemisfér. Tyto útvary jsou nejrozvinutější u živočichů s dobrým čichem a u čistě vodních forem jsou značně redukovány. Na základě rozdílů v jejich vývoji se rozlišuje několik typů struktury mozku. U vačnatců, hmyzožravců, bezzubých, masožravců, hlodavců a některých dalších jsou čichové cibule velké a dobře vyčnívají při pohledu na mozek shora. Tento typ mozku s dokonalým vyvinutím čichových laloků se nazývá makrosmatický. U ploutvonožců, sirén a mnoha primátů jsou cibule špatně vyvinuté; Tato zvířata se vyznačují mikrosmatickým mozkem. Konečně u kytovců tzv anosmatický mozek s redukovanými čichovými bulby. Dříve se mělo za to, že schopnost rozlišovat mezi chemickými signály byla u velryb a delfínů zcela ztracena, ale ukazuje se, že to není tak úplně pravda.
Povrchové vrstvy hemisfér předního mozku savců tvoří pallium neboli dřeňovou klenbu (pallium). Horní vrstva sestávající z těl neuronů a měkkých nervových vláken se nazývá kůra (cortex cerebri) a představuje šedou hmotu fornixu. Těla neuronů jsou umístěna v kůře ve vrstvách a tvoří zvláštní struktury obrazovky. Tato organizace mozku umožňuje prostorově reprezentovat vnější svět na základě informací přicházejících ze smyslů. Struktury obrazovek jsou charakteristické pro nejdůležitější mozková centra savců, zatímco u ostatních obratlovců jsou méně časté, hlavně ve zrakových centrech. Pod kůrou se nachází vrstva myelinizovaných neuronových výběžků – bílá hmota fornixu. Nervová vlákna bílé hmoty tvoří vodivé svazky, které pájejí hemisféry – mozkové komisury. I když je hlavní část šedé hmoty koncentrována v kůře, je přítomna i v podkorové vrstvě, kde je zastoupena relativně malými shluky – jádry. Patří mezi ně striatální tělíska (corpora striata), která leží pod dutinou postranní komory a dostala své jméno kvůli nervovým vláknům, které je protínají. Pod kontrolou kůry plní striatum funkci regulace stereotypních, automatizovaných reakcí – nepodmíněných reflexů.
Schémata odrážející progresivní vývoj hemisfér předního mozku. Boční pohled na polokouli s čichovou žárovkou. Různé řezy, které se od sebe liší cytologicky, se barví odlišně.
A - hemisféra je jen čichový lalok.
B - rozlišuje se dorzální úsek - archipallium (= hippocampus) a ventrální úsek - bazální jádro (striatum).
B - bazální ganglia se přesunula do vnitřní oblasti hemisféry.
D - objeví se malá oblast, představující neopallium.
D - paleopallium je vytlačeno na mediální plochu hemisféry, ale neopallium je stále skromné a pod čichovým sulkem jsou zachovány výrazně vyvinuté čichové úseky.
E - primitivní čichové oddělení je zachováno pouze ve ventrální oblasti a neopallium dosahuje extrémně silného vývoje. (Po Romer a Parsons, 1992.) U ryb se začala tvořit mozková klenba. Díky progresivnímu vývoji čichového smyslového systému se u nich vyvine paleopallium neboli starověký oblouk (paleopallium), zcela pokrývající malé hemisféry. Ve stádiu lalokoploutvých ryb se archipallium neboli starý oblouk (archipallium) objevuje v dorzální části hemisfér blíže k ose těla. U obojživelníků a primitivních plazů přijímá další vývoj, v důsledku čehož je paleopallium odsunuto stranou a je zachováno pouze podél bočního povrchu hemisfér. Paleopallium má přitom nadále převážně čichový charakter a ve vyšších fázích evoluce tvoří čichové laloky mozkové kůry. Archipallium je do jisté, i když malé míry korelačním centrem, přijímajícím vzestupná vlákna z diencefala, stejně jako vlákna z čichového bulbu a čichového laloku; zdá se také, že souvisí s emočním chováním. Nervová dráha z této oblasti do hypotalamu tvoří hlavní prvek svazku vláken nazývaného u savců fornix.
U obojživelníků se poprvé objevuje rudiment neopallia neboli nový oblouk (neopallium). U plazů už neopallium tvoří malou oblast mezi prastarou a starou klenbou. Od samého počátku svého vývoje je tato oblast asociativním centrem, podobně jako bazální ganglia, přijímající vlákna, která do ní přepínají senzorické signály z mozkového kmene a naopak předávají příkazy přímo motorickým sloupcům.
U monotremes je neopallium stále vklíněno mezi paleopallium zvenčí a archipalliem zevnitř. U vačnatců roste za střechu a boční stěny polokoulí. V tomto případě je archipallium přemístěno na mediální povrch a paleopallium je omezeno na ventrolaterální část hemisféry, která se nachází pod nosní rýhou (fissura rhinalis) - rýha, která představuje hranici mezi čichovou a nečichovou oblastí. kůry. V placentách v důsledku další komplikace a expanze neopallia dosáhnou hemisféry takové velikosti, že svým objemem převyšují ostatní části mozku dohromady. Hemisféry rostou dozadu a do stran a postupně pokrývají diencephalon, střední mozek a část mozečku. Párové komory a starověké struktury primárně zodpovědné za čich (čichové bulby, staré vrstvy dřeňové klenby a související nervové svazky a jádra) jsou v placentárním mozku odsunuty stranou a deformovány. Paleopallium je tedy zachováno na ventrálním povrchu hemisféry ve formě malé čichové oblasti zvané piriformní lalok (lobus piriformis) a archipallium je složeno hluboko do záhybu spánkového laloku do smotku zvaného hipokampus ( hippocampus). Převaha mozkových hemisfér nad ostatními částmi mozku je patrná u všech savců, zvláště výrazná je však u pokročilých forem, jako je člověk. Funkčně dominují i hemisféry. Na rozdíl od savců se komplikace předního mozku u ptáků projevují hlavně v růstu bazálních jader (nuclei basales), spíše než ve zbývajícím tenkém fornixu. 
Schematické řezy levou hemisférou předního mozku. Označení barev jsou stejná jako na předchozím obrázku.
1 - paleopallium; 2 - postranní komora; 3 - archipallium; 4 - bazální jádra; 5 - neopallium; 6 - corpus callosum.
A - primitivní stadium. Hemisféra je v podstatě čichový lalok. Špatně diferencovaná šedá hmota se nachází uvnitř mozku.
B - stadium pozorované u moderních obojživelníků. Šedá hmota se ještě nachází daleko od vnějšího povrchu, ale již se dělí na paleopallium (= čichový lalok), archipallium (= hippocampus) a bazální ganglia (= striatum). Ten nabývá významu asociativního centra, které má aferentní a eferentní spojení s thalamem (znázorněno čarami symbolizujícími nařezané svazky vláken).
B - progresivnější stádium, ve kterém jsou bazální ganglia ponořena do hemisféry, zatímco části kůry se posunuly poněkud ven.
G je stádium, ve kterém se nacházejí pokročilí plazi. Objeví se neopallium.
D - stadium primitivního savce. Neopallium se zvýšilo. Má rozsáhlé spojení s mozkovým kmenem. Archipalium zapnuto mediální povrch hemisféra se obaluje jako hipokampus. Paleopallium je stále vysoce vyvinuté.
E - stadium vysoce organizovaného savce. Neopallium extrémně roste a shromažďuje se v záhybech. Paleopallium zaujímá omezenou ventrální oblast, která představuje hruškovitý lalok. Rozvíjí se corpus callosum – mocný most spojující neopalliové oblasti obou hemisfér. (Po Romer a Parsons, 1992.) Kůra neopallia se nazývá neokortex nebo neokortex. U savců slouží jako centrum vyššího (podmíněného reflexu) nervová činnost, koordinující práci ostatních částí mozku. Odtud jsou impulsy posílány do různých orgánů a tkání těla a zde se v souladu s podmínkami prostředí provádí regulace fyziologických procesů. Právě nová kůra mozková hromadí stopy jednotlivých vzruchů a jejich kombinací, v důsledku čehož se obohacuje operační paměť poskytující možnost volit optimální řešení v nových situacích. Tato rozhodnutí častěji představují nové kombinace dříve známých prvků chování, ale také se vyvíjejí a konsolidují nové možnosti jednání. Nová kůra při svém vývoji přebírá nejen funkce korelačního a asociativního centra nově vznikajících typů vyšší nervové aktivity, ale začíná plnit i mnoho funkcí, které dříve patřily k centrům mozkového kmene a bazálních ganglií. V tomto případě nejsou prastará centra, která ovládají instinktivní činy, odstraněna, ale pouze podléhají vyšší kontrole.
V souvislosti s vývojem nového kortexu ztrácí střecha středního mozku svůj dřívější význam a zůstává pouze reflexním a přenosovým centrem. Sluchové a další somatické senzorické impulsy jsou přenášeny dopředu do thalamu, kde je přerušena většina zrakových vláken, a všechny tyto signály z thalamu jsou přenášeny do hemisfér podél silných nervových svazků. Podobná thalamická spojení s bazálními ganglii vznikla u málo organizovaných skupin obratlovců a byla nejvíce vyvinuta u ptáků. Na rozdíl od ptáků u savců většina vláken prochází striatem a rozchází se na povrch neokortexu. Tak k němu proudí kompletní soubor senzorických dat, na jejichž základě se v kůře dělají příslušná motorická „rozhodnutí“.
Jak již bylo zmíněno, některé signály jsou přenášeny z kůry do mozečku prostřednictvím mostu a zajišťují potřebné regulační účinky. Neokortex má také spojení se striatem a dokonce i s hypotalamem – a tedy s autonomním nervový systém. Převážná část motorických příkazů je však zasílána podél pyramidálního traktu (tractus corticospinalis) - speciálního nervového svazku, který přímo, bez přepínání, jde z mozkové kůry přes střední mozek do somatických motorických oblastí mozkového kmene a míchy. V tomto případě se vlákna laterální části této dráhy protínají a inervují opačnou stranu těla (tj. levé vlákno inervuje pravou část těla a naopak) a ventrální vlákna zůstávají spojena s jejich stranou. z těla. Pyramidový trakt je přítomen pouze u savců, což jasně dokládá dominantní postavení neokortexu u nich. Největší rozvoj tato struktura dosahuje u opic a zejména u lidí hraním důležitá role ve vzpřímeném držení těla. U vačnatců zasahují pyramidální axony pouze do hrudní oblasti, zatímco u monotremů pyramidální trakt zcela chybí. 
Komory lidského mozku; pohled z levé strany. Komory jsou znázorněny jako odlitek, ale mozková tkáň není zobrazena. S růstem hemisféry předního mozku se postranní komora rozšířila dozadu a vytvořila zadní roh v týlním laloku a v jeho boční části - dolů a dopředu, aby vytvořila postranní roh ve spánkovém laloku. Tyto výrůstky směřující dozadu a dolů vedly ke změnám v uspořádání různých částí mozku. Hipokampus, který se vyvinul v dorzální poloze na mediálním povrchu hemisféry, se u vysoce vyvinutých savců posunul dozadu a dolů do ventrální polohy. (Po Romer a Parsons, 1992.) Vzhledem k tomu, že neokortex je tenká vrstva vrstveného buněčného materiálu pod bílou vláknitou hmotou mozku, pouhé zvýšení objemu hemisfér nemůže způsobit úměrný růst kůry. Současně se v pokročilých formách může oblast kůry výrazně zvětšit díky jejímu skládání. Takto vytvořené záhyby se nazývají konvoluce (gyri) a hluboké mezery mezi nimi se nazývají drážky (sulci). Oba obsahují společné morfologické komponenty. V nejjednodušším případě je zde jedna hluboká Sylviova štěrbina oddělující čelní lalok (lobus frontalis) od spánkového laloku (lobus temporalis). Poté se nad a před Sylviovou trhlinou objeví příčně probíhající Rolandova trhlina, která odděluje frontální lalok od temenního laloku (lobus parietalis) nahoře. U primátů odděluje příčná rýha malý zadní týlní lalok (lobus occipitalis). Kromě hlavních rýh se tvoří mnoho dalších rýh; jejich počet je zvláště vysoký u primátů a zubatých velryb. Dříve se věřilo, že rýhy v některých případech označují morfologické hranice odpovídající určitým oblastem kůry. Další studie však ukázaly absenci pevného spojení mezi rozložením skládacích a konstrukční jednotka kůra (kromě nosní rýhy a do určité míry i centrální rýhy u primátů, o které bude řeč později). Je pozoruhodné, že kortikální skládání se u několika evolučních kmenů savců vyvinulo zcela nezávisle. U relativně primitivních savců, jako jsou monotreme, vačnatci, ale i někteří placentáři (hmyzožravci, chiropterani, hlodavci, zajícovci), je kůra skromněji vyvinutá a má hladký povrch. 
Umístění mozku v lebce fosilních a žijících psovitých šelem. Je patrný nárůst velikosti a složitosti mozku, zejména hemisfér předního mozku. Hesperocyon ( Hesperocyon gregarius) (vlevo) - oligocénní forma, která žila přibližně před 30 miliony let. Fennec ( Vulpes zerda) (napravo) - moderní forma podobné velikosti. (Po Romer a Parsons, 1992.) Šedá hmota neokortexu se vyznačuje složitou histologickou strukturou. U placentárních savců je 6 vrstev buněk ležících na sobě a mezi nimi uložených vláken; to značně odlišuje neokortex od zachovaných oblastí kůry paleopallia a archipallia, kde lze rozlišit pouze 2 až 4 vrstvy buněk. Současné odhady naznačují, že u savců, kteří mají zvláště velký mozek, může počet neokortikálních buněk dosáhnout miliard.
Bílá hmota, která se nachází pod šedou hmotou, kromě vějíře spojení vedoucího z kůry do spodních částí mozku a zpět, zahrnuje velké množství propletená příčná vlákna spojující různé oblasti samotné kůry. Takto vytvořená komisura je protažena dozadu (podle rychlosti růstu hemisfér) a je rozdělena na dvě desky srostlé podél zadního okraje. Spodní, tenčí a odkloněná směrem dolů předním okrajem, je fornix (fornix), komisura archipallium cortex (tj. hippocampus). Horní, tlustší horizontální komisura patří k nové kůře a nazývá se corpus callosum. Toto vzdělávání umožňuje spojit paměť obou hemisfér a výrazně zvyšuje schopnost mozku učit se. Corpus callosum je díky výraznému rozvoji neokortexu přítomno pouze v placentách, monotremům a vačnatcům chybí. Kromě toho mají všichni savci přední komisuru (commissura anterior), spojující čichové oblasti kůry. 
Vrstva po vrstvě uspořádání nervových buněk v kůře telencephalon savec (podle Naumova a Kartasheva, 1979.) Složitý systém„vodiče“ spojující všechny části kůry naznačuje, že šedá hmota je v principu jeden útvar, jehož všechny části mají stejné schopnosti vykonávat jakékoli funkce mozkových hemisfér. Do jisté míry to platí: experimenty ukazují, že u laboratorních zvířat je možné zničit významnou část neokortexu, aniž by došlo k trvalému narušení jejich normální činnosti. Údaje o úrazech a nemocnosti potvrzují, že to platí i pro lidský mozek. Zároveň je zřejmé, že určité oblasti kůry jsou běžně spojeny s výkonem velmi specifických funkcí. Výše byly zmíněny oblasti paleopallia a archipallia, určené hlavně pro analýzu čichových informací a zachované ve formě piriformního laloku a hipokampu. Diferenciace jednotlivých oblastí probíhá i v kortexu neopallia. Přední část hemisfér obsahuje motorickou oblast. Zde umístěný čelní lalok mimo jiné řídí komunikaci zvířat včetně akustické; u lidí je spojena s řečí, t. j. druhá bezpečnostní systém. Zadní strana hemisfér je spojena s vnímáním vjemů. Týlní lalok a temporální lalok obsahují oblasti, které kontrolují zrak a sluch. Dále vpřed, v blízkosti motorické oblasti, jsou oblasti, které přijímají hmatové a proprioceptivní signály. U primátů centrální sulcus (sulcus centralis), procházející vrcholem hemisféry od mediální k laterální ploše, vymezuje (i když ne zcela přesně) motorickou oblast od oblasti senzorické. Podél předního okraje centrálního sulku jsou v lineárním pořadí umístěny specifické motorické oblasti obsluhující každou část těla a končetiny. Podél zadního okraje centrálního sulku jsou ve stejném pořadí umístěny oblasti smyslového vnímání odpovídajících částí těla.
U mnoha savců je tedy téměř celý povrch neokortexu obsazen oblastmi více či méně úzce spojenými s určitými smyslovými nebo motorickými funkcemi. Ačkoli centrální sulcus může chybět, placenty obecně udržují podobné lineární uspořádání senzorických a motorických oblastí proti sobě. U vačnatců (a mezi placentami, u xenartridů) je „označení“ oblastí těla přibližně stejné, ale smyslové oblasti nejsou odděleny od motorických, ale střídají se s nimi. Ale například u lidí tyto specifické funkční oblasti zabírají relativně málo místa na povrchu neokortexu. Mezi nimi vznikly velké oblasti šedé hmoty (jedna zvlášť velká taková oblast zabírá většinu čelního laloku), které nejsou spojeny se specifickými senzorickými nebo motorickými funkcemi. Proto se těmto oblastem často říká „bílá místa“, i když, jak ukazuje poškození těchto oblastí, nacházejí se v nich naše vyšší mentální schopnosti, včetně učení, iniciativy, předvídavosti a úsudku. Zároveň existují i oblasti, které lze odstranit bez vážných následků pro intelektuální činnost. 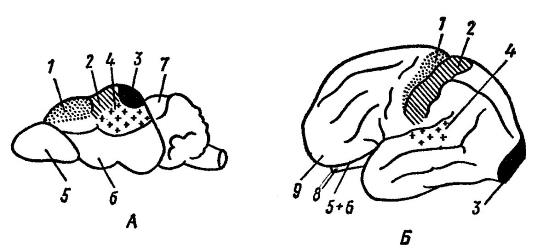
Funkční centra mozkové kůry rejska ( Sorex sp.) (A) a člověk ( Homo sapiens) (B) (podle Naumova a Kartasheva, 1979):
1 - motorický střed; 2 - centrum muskulokutánní citlivosti; 3 - zrakové centrum; 4 - sluchové centrum; 5 - čichová žárovka; 6 - čichové laloky; 7 - střecha středního mozku; 8 - mozeček; 9 - čelní lalok. O evoluci mozku velký vliv vyvíjené vnějším prostředím a motorickou (potravní, obrannou) činností. Vývoj různých částí mozku je přitom dán především metodami hledání potravy: u psa ( Canis lupus), při použití čichu v tomto procesu je čichová oblast rozvinutější; u kočky ( Felis silvestris), hledání potravy pomocí zraku – zrakového; u makaků ( Macaca mulatta), pomocí zraku a sluchu – zrakového a sluchového.
Obvykle se předpokládá, že velikost mozkových hemisfér určuje rozdíly v mentálních schopnostech různých savců. V v jistém smyslu to je pravda, ale se značnými výhradami. Větší mozek se skládá z více nervové buňky. Pokud dostupná plocha kortikálního povrchu nějakým způsobem souvisí s inteligencí, pak je jasné, že ze dvou mozkových variant stejné velikosti tím vyvinutější bude ten s rýhovaným povrchem a tím méně vyvinutý bude mozek s hladkým povrchem. Velikost samotného zvířete ovlivňuje i objem mozku. To se děje jen proto, že mozek musí mít větší oblasti, aby mohl sloužit rozsáhlejším senzorickým a motorickým spojením. Nárůst velikosti mozku však není zcela úměrný tělesné hmotnosti, takže velká zvířata mívají relativně menší mozky bez zjevné ztráty mentální kapacity. Absolutní velikost mozku tedy není absolutním kritériem inteligence. Nasvědčuje tomu jistě i fakt, že velrybí mozek může být objemově pětkrát větší než lidský mozek.
Srovnání mozků některých savců:
1 - kůň; 2 - pes; 3 - klokan; 4 - osoba; 5 - slon. Procento mozku v celkové tělesné hmotě se nazývá cefalizační index. U velkých hmyzožravců je to asi 0,6%, u malých - až 1,2%, u velkých kytovců - asi 0,3% au malých - až 1,7%. U většiny primátů je index cefalizace 1-2%. U lidí dosahuje 2-3 % a některé malé širokonosé opice mají mozek, jehož hmota tvoří až 7 % jejich tělesné hmotnosti. Současně se u moderních plazů a ptáků index cefalizace pohybuje od 0,05 do 0,5%.
Níže je uvedena hmotnost mozku některých savců (hmotnost zvířete je uvedena v závorkách):
Vačice virginská ( Didelphis virginiana) - 7,6 g (5 kg);
koala ( Phascolarctos cinereus) - 19,2 g (8 kg);
slon savana ( Loxodonta africana) - 6000 g (5000 kg);
ježek obecný ( Erinaceus europaeus) - 3,3 g (1 kg);
domácí myš ( Mus musculus) - 0,3 g (0,02 kg);
šedá krysa ( Rattus norvegicus) - 2 g (0,3 kg);
veverka obecná ( Sciurus vulgaris) - 7 g (0,4 kg);
Evropský králík ( Oryctolagus cuniculus- 11 g (3 kg);
domácí kůň ( Equus ferus) - 530 g (500 kg);
Černý nosorožec ( Diceros bicornis) - 500 g (1200 kg);
jelen běloocasý ( Odocoileus virginianus) - 500 g (200 kg);
žirafa ( Žirafa camelopardalis) - 680 g (800 kg);
domácí ovce ( Ovis orientalis) - 140 g (55 kg);
domácí býk ( Bos primigenius) - 490 g (700 kg);
Velbloud dvouhrbý ( Camelus bactrianus) - 762 g (700 kg);
hroch ( Hroch obojživelník) - 580 g (3500 kg);
delfín obecný ( Delphinus delphis) - 815 g (60 kg);
narval ( Monodon monoceros) - 2997 g (1578 kg);
vorvaň ( Physeter macrocephalus) - 8028 g (35833 kg);
modrá velryba ( Balaenoptera musculus) - 3636 g (50900 kg);
domácí kočka ( Felis silvestris) - 25 g (3 kg);
Lev ( Panthera leo) - 270 g (250 kg);
liška obecná ( Vulpes vulpes) - 53 g (4,5 kg);
domácí pes ( Canis lupus) - 64 g (10 kg);
lední medvěd ( Ursus maritimus) - 500 g (700 kg);
mrož ( Odobenus rosmarus) - 1130 g (700 kg);
kosman Geldi ( Callimico goeldii) - 7 g (0,2 kg);
kapucínka běločelá ( Cebus albifrons) - 57 g (1 kg);
makak rhesus ( Macaca mulatta) - 88 g (6,5 kg);
pavián ( Papio cynocephalus) - 200 g (25 kg);
stříbrný gibon ( Hylobates moloch) - 112 g (6,5 kg);
kalimantanský orangutan ( Pongo pygmaeus) - 413 g (50 kg);
západní gorila ( Gorila gorila) - 506 g (126 kg);
šimpanz obecný ( Pan troglodyty) - 430 g (55 kg);
rozumný člověk ( Homo sapiens) - 1400 g (72 kg).
Z výše uvedených příkladů je zřejmé, že u menších savců je mozek téměř vždy relativně větší, a jak se zvětšuje velikost zvířecího těla, relativní velikost mozku se zmenšuje. To je zvláště výrazné u blízce příbuzných druhů savců – například u koček ( Felis silvestris) a lev ( Panthera leo). Psi jsou v tomto smyslu také velmi přesvědčiví. různá plemena. Pokud jsou tělesné hmotnosti nejmenšího a největšího plemene přibližně v poměru 1:33, pak jsou mozkové hmoty stejných plemen v poměru 1:3. 
Rozsahy hodnot hmotnosti mozku a těla pro některé skupiny obratlovců. U domestikovaných zvířat, zbavených potřeby získávat potravu a bránit se nepřátelům, se výrazně zmenšuje velikost mozku. Například objem vlčího mozku ( Canis lupus) o 30 % více než u psa stejné velikosti. Zajímavé je, že tyto změny se týkají nejen tradičně domácích zvířat, ale i zástupců volně žijících druhů chovaných nějakou dobu v zajetí. Ano, lišky ( Vulpes vulpes), narozeni v přírodě, ale od prvních dnů žijících v zajetí mají menší mozek než jejich příbuzní žijící v přírodních podmínkách. V tomto případě rozdíly dosahují 20 %, což přibližně odpovídá rozdílu v objemu mozku divokých a skutečných domácích zvířat. Zmenšení mozku, i když ne tak výrazné (přibližně 5 %), bylo zjištěno u vlků chovaných v zajetí ( Canis), fretky ( Mustela), krysy ( Rattus). V tomto případě pokles neovlivňuje všechny části mozku, ale pouze ty oblasti, které jsou spojeny s fungováním smyslových orgánů. Nejpozoruhodnější je, že se zvyšuje hmotnost mozku vypuštěných domácích zvířat. Například divoké kočky mají mozek asi o 10 % větší než jejich domestikované protějšky. Významné zvětšení mozku bylo také zjištěno u divokých králíků ( Oryctolagus cuniculus) na Kerguelenských ostrovech. Divocí osli ( Equus asinus) V Jižní Amerika měl 15 % velký mozek než ty domácí. Je také zajímavé, že mozek neandrtálce ( Homo neanderthalensis) a paleolitický homo sapiens ( Homo sapiens) byly o něco větší než mozek moderních lidí.
Bylo zjištěno, že mnoho savců vykazuje motorickou asymetrii, tj. převládající používání pravé nebo levé poloviny těla. Například při studiu nezlomených koní ( Equus ferus) zaznamenal, jakou nohou zvířata začínají chodit, na kterou stranu obcházejí překážky nejraději a na kterou stranu raději leží ve stáji na seně. V důsledku toho byla většina klisen praváků a většina hřebců leváků. Přibližně 10 % koní nevykazovalo žádnou preferenci pravé ani levé končetiny. Podle pozorování asi 90 % mrožů ( Odobenus rosmarus) vyhrabat měkkýše z mořského bahna pomocí správné ploutve. Kolébá mláďata, asi 80 % samic šimpanzů ( Pánev) a gorily ( Gorila) přitisknou hlavu k levé straně hrudníku (u žen je pozorováno přibližně stejné procento). Krysy ( Rattus), hledání potravy pomocí vibrissae umístěných na pravá strana tlamy, jsou větší kořistí než jejich levorucí příbuzní.
textová_pole
textová_pole
arrow_upward
Diencephalon je spolu s mozkovým kmenem pokryt shora a ze stran velké polokoule - telencephalon. Hemisféry se skládají z podkorových ganglií (bazálních ganglií) a mají dutiny -. Vnější strana polokoulí je pokryta (pláštěm).
Bazální ganglia nebo subkortikální ganglia
textová_pole
textová_pole
arrow_upward
Bazální ganglia nebo podkorové uzliny (nuclei basales)– útvary jsou fylogeneticky starší než kůra. Bazální ganglia dostala své jméno díky tomu, že leží na bázi mozkových hemisfér, v jejich bazální části. Patří mezi ně kaudátní a čočkovitá jádra, spojená ve striatu, plotu a amygdale.
Kádové jádro
textová_pole
textová_pole
arrow_upward
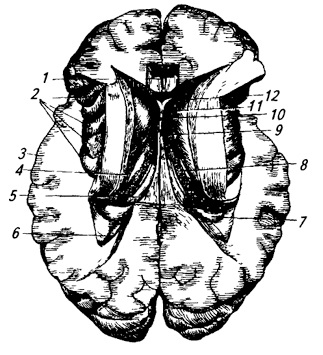
Kádové jádro (nucleus caudatus) protáhlé v sagitální rovině a silně zakřivené (obr. 3.22; 3.32; 3.33). Jeho přední, zesílená část je hlava– nachází se před optickým thalamem, v laterální stěně předního rohu postranní komory, za ním se postupně zužuje a přechází v ocas. Caudate nucleus pokrývá zrakový talamus vpředu, nahoře a po stranách.
Rýže. 3.22.
1 – nucleus caudatus;
2 – sloupy klenby;
3 – epifýza;
4 – horní a
5 – colliculus inferior;
6 – vlákna středního cerebelárního peduncle;
7 – dráha horního cerebelárního peduncle (připravená);
8 – jádro stanu;
9 – červ;
10 – kulový,
11 – korkové a
13 – zubaté jádro;
12 – kůra mozečku;
14 – horní cerebelární stopka;
15 – vodítko trojúhelník;
16 – talamický polštář;
17 – zrakový talamus;
18 – zadní komisura;
19 – třetí komora;
20 – přední jádro zrakového thalamu
 Rýže. 3.32.
Rýže. 3.32. Rýže. 3.32. Mozek - horizontální řez postranními komorami:
1 – corpus callosum;
2 – ostrov;
3 – kůra;
4 – ocas caudatus nucleus;
5 – klenba;
6 – zadní roh postranní komory;
7 – hipokampus;
8 – plexus choroideus;
9 – foramen interventrikulární;
10 – průhledná přepážka;
11 – hlava nucleus caudatus;
12 – přední roh postranní komory
Jádro ve tvaru čočky
textová_pole
textová_pole
arrow_upward
Jádro ve tvaru čočky (nucleus lentiformis) nachází se mimo optický thalamus, na úrovni insula. Tvar jádra se blíží trojúhelníkovému jehlanu, jehož základna směřuje ven. Jádro je zřetelně rozděleno vrstvami bílé hmoty na tmavší boční část - skořápka a mediální - světlá koule, skládající se ze dvou segmentů: vnitřního a vnějšího (obr. 3.33; 3.34).
 Rýže. 3.33.
Rýže. 3.33. Rýže. 3.33. Horizontální řez mozkovými hemisférami na úrovni bazálních ganglií:
1 - corpus callosum;
2 – klenba;
3 – přední roh postranní komory;
4 – hlava nucleus caudatus;
5 – vnitřní kapsle;
6 – plášť;
7 – globus pallidus;
8 – vnější kapsle;
9 – plot;
10 – thalamus;
11 – epifýza;
12 – ocas caudatus nucleus;
13 – choroidální plexus postranní komory;
14 – zadní roh postranní komory;
15 – cerebelární vermis;
16 – quadrigeminální;
17 – zadní komisura;
18 – dutina třetí komory;
19 – jáma boční drážky;
20 – ostrov;
21 – přední komisura
 Rýže. 3.34.
Rýže. 3.34. Rýže. 3.34. Frontální řez mozkovými hemisférami na úrovni bazálních ganglií:
1 - corpus callosum;
2 – postranní komora;
3 – nucleus caudate (hlavička);
4 – vnitřní kapsle;
5 - jádro čočkovitého tvaru;
6 – boční drážka;
7 - temporální lalok;
8 – plot;
9 – ostrov;
10 – vnější kapsle;
11 – průhledná přepážka;
12 – vyzařování corpus callosum;
13 – mozková kůra
Shell
textová_pole
textová_pole
arrow_upward
 Rýže. 3.35.
Rýže. 3.35. Shell (putamen) podle genetických, strukturálních a funkčních charakteristik se blíží nucleus caudatus.
Oba tyto útvary mají složitější strukturu než globus pallidus. Vlákna se k nim přibližují především z mozkové kůry a thalamu (obr. 3.35).
Rýže. 3.35. Aferentní a eferentní spojení bazálních ganglií:
1 - precentrální gyrus;
2 – skořepina;
3 – vnější a vnitřní segmenty globus pallidus;
4 – čočkovitá smyčka;
5 - retikulární formace;
6 – retikulospinální trakt,
7 - rubrospinální trakt;
8 – cerebellotalamický trakt (z dentatus nucleus cerebellum);
9 – červené jádro;
10 – substantia nigra;
11 – subtalamické jádro;
12 – Zona incerta;
13 – hypotalamus;
14 – ventrolaterální,
15 – intralaminární a centromedická jádra thalamu;
16 – III komora;
17 – nucleus caudatus
Bledý míč
textová_pole
textová_pole
arrow_upward
Globus pallidus (globus pallidus) je spojen především s vedením vzruchů po četných sestupných drahách do podložních struktur mozku – červené jádro, substantia nigra atd. Vlákna z neuronů globus pallidus směřují do stejných jader mozku. thalamus, které jsou spojeny s mozečkem. Z těchto jader jdou četné cesty do mozkové kůry.
Globus pallidus přijímá impulsy z caudatus nucleus a putamen.
Striatum (corpus striatum), které spojuje caudatum a lentiformní jádro, patří k eferentním extrapyramidový systém. Dendrity striatálních neuronů jsou pokryty četnými trny. Na nich končí vlákna z neuronů kůry, thalamu a substantia nigra (obr. 3.35). Striatální neurony zase posílají axony do intralaminárních, předních a laterálních jader thalamu. Z nich jdou vlákna do kortexu a tím se uzavře zpětnovazební smyčka mezi korovými neurony a striatem.
Během procesu fylogeneze byla tato jádra postavena na jádrech středního mozku. Striatum, které přijímá impulsy z thalamu, se účastní tak složitých automatických pohybů, jako je chůze, lezení a běh. V jádrech striata jsou uzavřeny oblouky nejsložitějších nepodmíněných, tzn. vrozené reflexy. Extrapyramidový systém je fylogeneticky starší než pyramidový systém. U novorozence tato ještě není dostatečně vyvinutá a impulsy do svalů jsou dodávány z podkorových ganglií přes extrapyramidový systém. V důsledku toho jsou pohyby dítěte v prvních měsících života charakterizovány generalizací a nediferenciací. Jak se mozková kůra vyvíjí, axony jejich buněk přirůstají k bazálním gangliím a jejich činnost začíná být řízena kůrou. Subkortikální ganglia jsou spojena nejen s motorickými reakcemi, ale také s autonomními funkcemi - ty jsou nejvyšší subkortikálních center autonomní nervový systém.
Amygdala
textová_pole
textová_pole
arrow_upward
Amygdala (corpus atugdaloideum) (amygdala) – soubor buněk v bílé hmotě spánkového laloku. S pomocí přední komisura spojuje se se stejnojmenným tělem na druhé straně. Amygdala přijímá impulsy z různých aferentních systémů, včetně čichového systému, a souvisí s emocionální reakce(obr. 3.36).
 Rýže. 3.36.
Rýže. 3.36. Rýže. 3.36. Mozkové struktury spojené s amygdalou: aferentní (A) a eferentní (B) spojení amygdaly:
1 - thalamická jádra;
2 – periakveduktální šedá hmota;
3 – parabrachiální jádro;
4 – modrá skvrna;
5 - jádra stehů;
6 – jádro osamělého traktu;
7 - dosální jádro n. X;
8 – spánková kůra;
9 – čichová kůra;
10 – čichová žárovka;
11 - čelní kůra;
12 – gyrus cingulární;
13 – corpus callosum;
14 – čichové jádro;
15 - anteroventrální a
16 – dorzomediální jádro thalamu;
17 – centrální,
18 – kortikální a
19 – bazolaterální jádro amygdaly;
20 – hypotalamus;
21 – retikulární formace;
22 – přepážka;
23 – substantia nigra;
24 – ventromediální jádro hypotalamu; XXIII, XXIV, XXVIII – korová pole
>>Funkce předního mozku
§ 46. Funkce předního mozku
1. Kde se rozpoznávají vnímané obrazy?
2. Plní levá a pravá hemisféra stejné funkce?
Přední mozek se skládá ze dvou částí: diencephalon a mozkové hemisféry mozek. Jedná se o největší část mozku, která se skládá z pravé a levé poloviny.
Diencephalon se skládá ze tří částí – horní, střední a dolní (obr. 93-97). Centrální část diencephalonu se nazývá thalamus. Skládá se ze dvou párových útvarů oddělených třetí mozkovou komorou. Všichni se sem hrnou informace ze smyslů. Zde přichází první hodnocení jeho významu. Jen díky thalamu důležitá informace vstupuje do mozkové kůry.
Spodní část diencefala se nazývá hypotalamus. Reguluje to metabolismus a energie. V jeho jádrech jsou centra žízně a jejího utišení, hladu a nasycení. Hypotalamus řídí uspokojování potřeb a udržování stálého vnitřního prostředí – homeostázy. Za účasti diencephalonu a dalších částí mozku se provádí mnoho cyklických pohybů: chůze, běh, skákání, plavání atd., jakož i udržování polohy mezi pohyby.
Mozkové hemisféry jsou rozděleny hlubokou předozadní trhlinou na levou a pravou část. V jeho hlubinách je spojuje most bílé hmoty - corpus callosum.
Povrch velkého mozku je tvořen kůrou, tvořenou šedou hmotou. Jsou tam soustředěna těla neuronů. Jsou uspořádány do sloupců, které tvoří několik vrstev.
Kůra je bílá látka skládající se z hmoty nervových vláken spojující kortikální neurony mezi sebou a se spodními částmi mozku. V tloušťce hemisfér se mezi bílou hmotou nacházejí ostrůvky šedé hmoty ve formě jader, tvořících podkorová centra.
Povrch polokoulí je složený. Vyčnívající části povrchu tvoří záhyby a prohlubně tvoří drážky. Výrazně zvětšují povrch mozkové kůry. Nejhlubší rýhy rozdělují každou hemisféru na čtyři laloky: čelní, parietální, okcipitální a temporální (obr. 95). Sousedí s odpovídajícími kostmi, a proto nesou jejich jména. Centrální sulcus odděluje frontální lalok od parietálního laloku, laterální sulcus odděluje temporální lalok od frontálního a parietálního laloku.
Analýza probíhá v neuronech mozkové kůry nervové vzruchy, vycházející ze smyslů (obr. 96). Provádí se v citlivých oblastech, které zabírají střední a zadní část mozku. Neurony jsou tedy soustředěny v okcipitálním laloku vizuální oblast, v časovém - sluchovém. V parietální zóně za centrálním gyrusem je oblast muskulokutánní citlivosti.
Čichové a chuťové zóny se nacházejí na vnitřním povrchu spánkových laloků. Centra, která regulují aktivní chování, se nacházejí v předních částech mozku, ve frontálních lalocích mozkové kůry.
Motorická zóna se nachází před centrálním gyrusem.
Pravá hemisféra ovládá orgány levé strany těla a přijímá informace z prostoru vlevo. Levá hemisféra reguluje fungování orgánů pravé strany těla a vpravo vnímá informace z prostoru.

Hlavním rysem velkého lidského mozku je to, že pravá a levá hemisféra jsou funkčně odlišné. V levé hemisféře mají praváci zpravidla centra řeči. Zde se analyzuje situace a související akce podle jednotlivých parametrů, vyvíjejí se zobecnění a vyvozují se logické závěry. Pravá hemisféra vnímá situaci jako celek. Zde vznikají takzvaná intuitivní řešení. Pravá hemisféra rozpoznává obrázky a melodie a pamatuje si tváře.
V mozkových hemisférách se vytvářejí dočasná spojení mezi signálem, podmíněnými reflexními podněty a vitálními událostmi. Prostřednictvím těchto spojení se shromažďují individuální zkušenosti.
Stará a nová mozková kůra. Stará kůra je již přítomna u plazů. U savců je jeho vzhled spojen s rozvojem čichu. Jako pás obklopuje základ mozku a zahrnuje subkortikální jádra (obr. 97).
Jsou zde soustředěna centra spojená se složitými instinkty, emocemi a pamětí. Stará kůra umožňuje tělu rozlišovat příznivé a nepříznivé události a reagovat na ně strachem, radostí, agresí a úzkostí. Zde se do paměti ukládají informace o prožitých událostech. To umožňuje za podobných okolností podnikat kroky, které povedou k úspěchu. Na rozdíl od neokortexu nedokáže stará kůra přesně rozpoznat objekty, odhadnout pravděpodobnost budoucích událostí a plánovat reakce na jejich výskyt.
A nová kůra přijímá informace z vnitřních orgánů a ze smyslových orgánů. Ve frontálních lalocích se z četných potřeb vybere ta nejdůležitější a vytvoří se cíl činnosti, plán k dosažení cíle na základě rozboru situace a minulých zkušeností.
Zde se za účasti řečových center rozvíjejí scénáře budoucího chování. Realizují je další části mozku a míchy spojené s výkonnými orgány.

Informace o dosažených výsledcích přicházejí prostřednictvím zpětná vazba PROTI čelní laloky hemisféry a v závislosti na dosaženém účinku se činnost zastaví nebo pokračuje v modifikované podobě.
Přední mozek; diencephalon: thalamus, hypothalamus; mozkové hemisféry, corpus callosum, kortex, sulci, gyri, mozkové laloky: senzorické a motorické oblasti; dočasné (podmíněné reflexní) spojení; stará a nová kůra.
1. Jaká oddělení se rozlišují v předním mozku?
2. Jaké jsou funkce thalamu a hypotalamu?
3. Proč je povrch polokoulí složený?
4. Jak je distribuována šedá a bílá hmota v mozkových hemisférách? Jaké funkce plní?
5. Jaké jsou funkce staré kůry mozkové?
6. Jak jsou distribuovány funkce mezi levou a pravou hemisférou velkého mozku?
7. Které spoje v těle se nazývají přímé a které reverzní?
Vysvětlete, ve které hemisféře došlo k poškození motorických center, pokud pacient zažil ochrnutí pravé nohy nebo paže.
Kolosov D.V. Mash R.D., Belyaev I.N. Biologie 8. tř.
Odeslali čtenáři z webu



