Строение больших полушарий головного мозга человека. Строение и значение больших полушарий головного мозга
Общий обзор строения больших полушарий
Большие полушария головного мозга представляют собой самый массивный отдел головного мозга. Они покрывают мозжечок и ствол мозга. Большие полушария составляют примерно 78% от общей массы мозга. В процессе онтогенетического развития организма большие полушария головного мозга развиваются из коечного мозгового пузыря нервной трубки, поэтому данный отдел головного мозга называется также конечным мозгом.
Большие полушария головного мозга разделены по средней линии глубокой вертикальной щелью на правое и левое полушария.
В глубине средней части оба полушария соединены между собой большой спайкой – мозолистым телом. В каждом полушарии различают доли; лобную, теменную, височную, затылочную и островок.
Доли мозговых полушарий отделяются одна от другой глубокими бороздами. Наиболее важны три глубокие борозды: центральная (роландова) отделяющая лобную долю от теменной, боковая (сильвиева) отделяющая височную долю от теменной, теменно-затылочная отделяющая теменную долю от затылочной на внутренней поверхности полушария.
Каждое полушарие имеет верхнебоковую (выпуклую), нижнюю и внутреннюю поверхность.
Каждая доля полушария имеет мозговые извилины, отделенные друг от друга бороздами. Сверху полушарие покрыто корой ~ тонким слоем серого вещества, которое состоит из нервных клеток.
Кора головного мозга – наиболее молодое в эволюционном отношении образование центральной нервной системы. У человека она достигает наивысшего развития. Кора головного мозга имеет огромное значение в регуляции жизнедеятельности организма, в осуществлении сложных форм поведения и становлении нервно-психических функций.
Под корой находится белое вещество полушарий, оно состоит из отростков нервных клеток – проводников. Из-за образования мозговых извилин общая поверхность коры головного мозга значительно увеличивается. Общая площадь коры полушарий составляет 1200 см 2 , причем 2/3 ее поверхности находится в глубине борозд, а 1/3 – на видимой поверхности полушарий. Каждая доля мозга имеет различное функциональное значение.
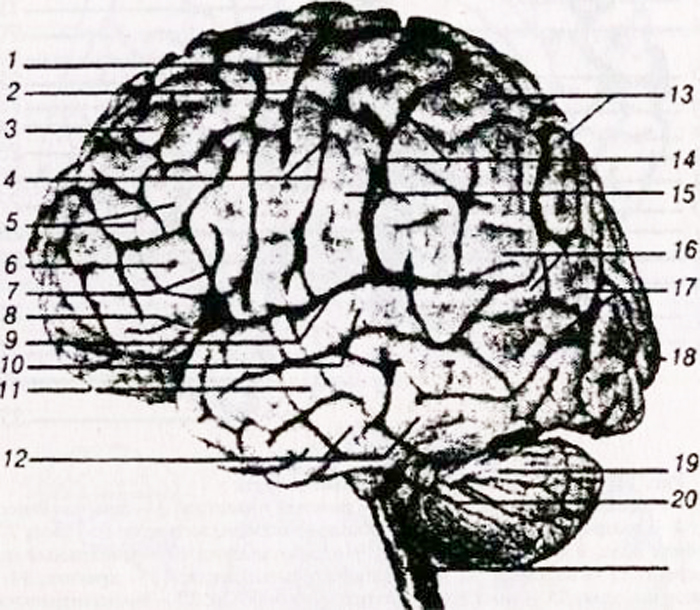
а – верхнебоковая поверхность: 1 – нижняя лобная извилина; 2 – средняя лобная извилина; 3 – верхняя лобная извилина; 4 – передняя центральная извилина; 5 – центральная (роландова) борозда; 6 – задняя центральная извилина; 7 – верхняя теменная долька; 8 – нижняя теменная долька; 9 – надкраевая (супрамаргинальная) борозда; 10 – угловая (ангулярная) борозда; 11 – теменно-затылочная борозда; 12 – нижняя височная извилина;
13 – средняя височная извилина; 14 – верхняя височная извилина; 15 – боковая (сильвиева) борозда;
б – внутренняя поверхность: 1 парацентральная долька; 2 – центральная борозда; 3 – поясная извилина: 4 – мозолистое тело; 5 – теменно-затылочная борозда; 6 – клин; 7 – шпорная борозда; 8 – язычковая извилина; 9 – извилина гиппокампа (парагиппокамповая извилина).
Строение лобной доли
Лобная доля занимает передние отделы полушарий. От теменной доли она отделяется центральной бороздой, от височной – боковой бороздой. В лобной доле имеются четыре извилины: одна вертикальная – прецентральная и три горизонтальные – верхняя, средняя и нижняя лобные извилины. Извилины отделены друг от друга бороздами.
На нижней поверхности лобных долей различают прямую и орбитальную извилины. Прямая извилина залегает между внутренним краем полушария, обонятельной бороздой и наружным краем полушария.
В глубине обонятельной борозды лежат обонятельная луковица и обонятельный тракт.
Лобная доля человека составляет 25–28% коры; средняя масса лобной доли 450г.
Функция лобных долей связана с организацией произвольных движений, двигательных механизмов речи, регуляцией сложных форм поведения, процессов мышления. В извилинах лобной доли сконцентрировано несколько функционально важных центров. Передняя центральная извилина является «представительством» первичной двигательной зоны со строго определенной проекцией участков тела. Лицо «расположено» в нижней трети извилины, Рука – в средней трети, нога – в верхней трети. Туловище представлено в задних отделах верхней лобной извилины. Таким образом, человек спроецирован в передней центральной извилине вверх ногами и вниз головой.
Передняя центральная извилина вместе с прилегающими задними и отделами лобных извилин выполняет очень важную в функциональном отношении роль. Она является центром произвольных движений. В глубине коры центральной извилины от так называемых пирамидных клеток – центрального двигательного нейрона – начинается основной двигательный путь – пирамидный, кортикоспинальный, путь. Периферические отростки двигательных нейронов выходят из коры, собираются в единый мощный пучок, проходят центральное белое вещество полушарий и через внутреннюю капсулу входят в ствол мозга; в конце ствола мозга они частично перекрещиваются (переходя с одной стороны на другую) и затем спускаются в спинной мозг. Эти отростки заканчиваются в сером веществе спинного мозга. Там они вступают в контакт с периферическим двигательным нейроном и передают ему импульсы из центрального двигательного нейрона. По пирамидному пути передаются импульсы произвольного движения.
В задних отделах верхней лобной извилины располагается также экстрапирамидный центр коры, тесно связанный анатомически и функционально с образованиями так называемой экстрапирамидной системы. Экстрапирамидная система – двигательная система, помогающая осуществлению произвольного движения. Это система «обеспечения» произвольных движений. Будучи филогенетически более старой, экстрапирамидная система у человека обеспечивает автоматическую регуляцию «заученных» двигательных актов, поддержание общего мышечного тонуса, готовность периферического двигательного аппарата к совершению движений, перераспределение мышечного тонуса при движениях. Кроме того, она участвует в поддержании нормальной позы.
Двигательные зоны коры
находятся в основном в
прецентральной извилине (поля 4 и 6) и парацентральной дольке на медиальной
поверхности полушария. Выделяют первичную и вторичную области – поля 4 и 6. Эти
поля моторные, но по своей характеристике, согласно исследованиям Института
мозга, они разные. В первичноймоторной
коре
(поле 4) расположены нейроны, иннервирующие мотонейроны мышц
лица, туловища и конечностей.
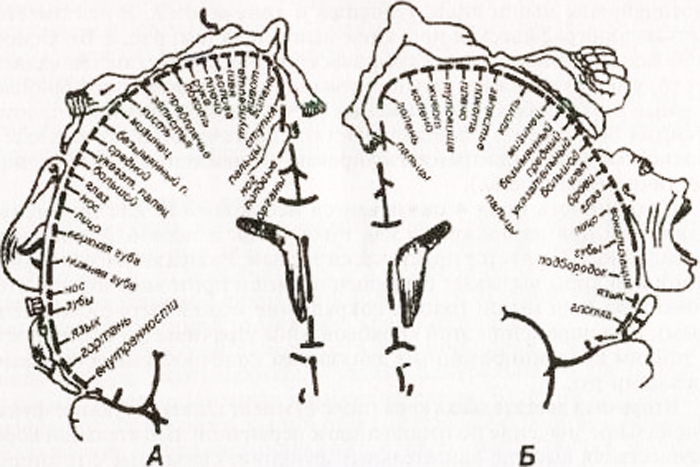
Схема самототопической проекции общей чувствительности и двигательных функций в коре головного мозга (по У. Пенфилду):
А – корковая проекция общей чувствительности; Б – корковая проекция двигательной системы. Относительные размеры органов отражают ту площадь коры головного мозга, с которой могут быть вызваны соответствующие ощущения и движения
В ней имеется четкая топографическая проекция мышц тела. Основной закономерностью топографического представительства является то, что регуляция деятельности мышц, обеспечивающих наиболее точные и разнообразные движения (речь, письмо, мимика), требует участия больших по площади участков двигательной коры. Поле 4 полностью занято центрами изолированных движений, поле 6 – лишь частично (подполе 6а).
Сохранность поля 4 оказывается необходимой для получения движений при раздражении как поля 4, так и поля 6. У новорожденного поле 4 является практически зрелым. Раздражение первичной моторной коры вызывает сокращение мышц противоположной стороны тела (для мышц головы сокращение может быть билатеральным). При поражении этой корковой зоны утрачивается способность к тонким координированным движениям конечностями и особенно пальцами рук.
Вторичная двигательная кора (поле 6) имеет главенствующее функциональное значение по отношению к первичной двигательной коре, осуществляя высшие двигательные функции, связанные с планированием и координацией произвольных движений. Здесь в наибольшей степени регистрируется медленно нарастающий отрицательный потенциал готовности, возникающий примерно за 1 с до начала движения. Кора поля 6 получает основную часть импульсации от базальных ганглиев и мозжечка, участвует в перекодировании информации о сложных движениях.
Раздражение коры поля 6 вызывает сложные координированные движения, например поворот головы, глаз и туловища в противоположную сторону, содружественные сокращения сгибателей или разгибателей на противоположной стороне. В премоторной коре расположены двигательные центры, связанные с социальными функциями человека: центр письменной речи в заднем отделе средней лобной извилины (поле 6), центр моторной речи Брока в заднем отделе нижней лобной извилины (поле 44), обеспечивающие речевой, а также музыкальный моторный центр (поле 45), обеспечивающий тональность речи, способность петь. Нижняя часть поля б (подполе бор), расположенная в области покрышки, реагирует на электроток ритмическими жевательными движениями. Нейроны двигательной коры получают афферентные входы через таламус от мышечных, суставных и кожных рецепторов, от базальных ганглиев и мозжечка. Основным эфферентным выходом двигательной коры на стволовые и спинальные моторные центры являются пирамидные клетки V слоя.
В заднем отделе средней лобной извилины находится лобный глазодвигательный центр, осуществляющий контроль за содружественным, одновременным поворотом головы и глаз (центр поворота головы и глаз в противоположную сторону). Раздражение этого центра вызывает поворот головы и глаз в противоположную сторону. Функция этого центра имеет огромное значение в осуществлении так называемых ориентировочных рефлексов (или рефлексов «что такое?»), имеющих очень важное значение для сохранения жизни животных.
Лобный отдел коры больших полушарий принимает также активное участие в формировании мышления, организации целенаправленной деятельности, перспективном планировании.
Строение теменной доли
Теменная доля занимает верхнебоковые поверхности полушария. От лобной теменная доля спереди и сбоку ограничивается центральной бороздой, от височной снизу – боковой бороздой, от затылочной – воображаемой линией, проходящей от верхнего края теменно-затылочной борозды до нижнего края полушария.
На верхнебоковой поверхности теменной доли имеются три извилины: одна вертикальная – задняя центральная и две горизонтальные – верхнетеменная и нижнетеменная. Часть нижнетеменной извилины, огибающей задний отдел латеральной борозды, называют надкраевой (супрамаргинальной), часть, окружающую верхнюю височную извилину, – узловой (ангулярной) областью.
Теменная доля, как и лобная, составляет значительную часть полушарий головного мозга. В филогенетическом отношении в ней выделяют старый отдел – заднюю центральную извилину, новый – верхнетеменную извилину и более новый – нижнетеменную извилину.
Функция теменной доли связана с восприятием и анализом чувствительных раздражений, пространственной ориентацией. В извилинах теменной доли сконцентрировано несколько функциональных центров.
В задней центральной извилине спроецированы центры чувствительности с проекцией тела, аналогичной таковой в передней центральной извилине. В нижней трети извилины спроецировано лицо, в средней трети – рука, туловище, в верхней трети – нога. В верхней теменной извилине находятся центры, ведающие сложными видами глубокой чувствительности: мышечно-суставным, двухмерно-пространственным чувством, чувством веса и объема движения, чувством распознавания предметов на ощупь.
Кзади от верхних отделов задней центральной извилины локализуется центр, обеспечивающий способность узнавания собственного тела, его частей, их пропорций и взаимоположение (поле 7).
Поля 1, 2, 3 постцентральной области составляют основное корковое ядро кожного анализатора. Вместе с полем 1 поле 3 является первичной, а поле 2 – вторичной проекционной зоной кожного. анализатора. Эфферентными волокнами постцентральная область связана с подкорковыми и стволовыми образованиями, с прецентральной и другими областями коры большого мозга. Таким образом, в теменной доле локализуется корковый отдел чувствительного анализатора.
Первичные сенсорные зоны – это области сенсорной коры, раздражение или разрушение которых вызывает четкие и постоянные изменения чувствительности организма (ядра анализаторов, по И.П. Павлову). Они состоят главным образом из мономодальных нейронов и формируют ощущения одного качества. В первичных сенсорных зонах обычно имеется четкое пространственное (топографическое) представительство частей тела, их рецепторных полей.
Вокруг первичных сенсорных зон находятся менее локализованные вторичные сенсорные зоны, нейроны которых отвечают на действие нескольких раздражителей, т.е. они полимодальны.
Важнейшей сенсорной областью являются теменная кора постцентральной извилины и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий, которую обозначают как соматосенсорная область I. Здесь имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, Инте-роцептивной чувствительности и чувствительности опорно-двигательного аппарата – от мышечных, суставных, сухожильных рецепторов.
Кроме соматосенсорной области I выделяют меньших размеров соматосенсорную область II, расположенную на границе пересечения центральной борозды с верхним краем височной доли, в глубине латеральной борозды. Степень локализации частей тела здесь выражена в меньшей степени.
В нижней теменной доле расположены центры праксиса. Под праксисом понимаются ставшие автоматизированными в процессе повторений и упражнений целенаправленные движения, которые вырабатываются в процессе обучения и постоянной практики в течение индивидуальной жизни. Ходьба, еда, одевание, механический элемент письма, различные виды трудовой деятельности (например, движения водителя по управлению автомобилем, косьба и пр.) являются праксисом. Праксис – высшее проявление свойственной человеку двигательной функции. Он осуществляется в результате сочетанной деятельности различных территорий коры головного мозга.
В нижних отделах передней и задней центральных извилин располагается центр анализатора интероцептивных импульсов внутренних органов и сосудов. Центр имеет тесные связи с подкорковыми вегетативными образованиями.
Строение височной доли
Височная доля занимает нижнебоковую поверхность полушарий. От лобной и теменной долей височная доля ограничиваете боковой бороздой. На верхнебоковой поверхности височной доле имеются три извилины: верхняя, средняя и нижняя.
Верхняя височная извилина находится между сильвиевой и верхней височной бороздами, средняя – между верхней и нижней височной бороздами, нижняя – между нижней височной бороздой и поперечной мозговой щелью. На нижней поверхности височной доли различают нижнюю височную извилину, боковую затылочно-височную извилину, извилины гиппокампа (ноги морского коня).
Функция височной доли связана с восприятием слуховых, вкусовых, обонятельных ощущений, анализом и синтезом речевых звуков, механизмами памяти. Основной функциональный центр верхнебоковой поверхности височной доли находится в верхней височной извилине. Здесь располагается слуховой, или гностический, центр речи (центр Вернике).
Хорошо изученной первичной проекционной зоной является слуховая кора (поля 41, 42, 52), которая расположена в глубине латеральной борозды (кора поперечных височных извилин Гешля). К проекционной коре височной доли относится также центр вестибулярного анализатора ● в верхней и средней височных извилинах (поля 20 и 21).
Обонятельная проекционная область находится в гиппокамповой извилине, особенно в ее переднем отделе (так называемом крючке). Рядом с обонятельными проекционными зонами находятся вкусовые.
Височные доли играют важную роль в организации сложных психических процессов, в частности памяти.
Cтроение затылочной доли
Затылочная доля занимает задние отделы полушарий. На выпуклой поверхности полушария затылочная доля не имеет резких границ, отделяющих ее от теменной и височной долей, за исключением верхнего отдела теменно-затылочной борозды, которая, располагаясь на внутренней поверхности полушария, отделяет теменную долю от затылочной. Борозды и извилины верхнебокой поверхности затылочной доли непостоянны и имеют вариабельное строение. На внутренней поверхности затылочной доли имеется шпорная борозда, которая отделяет клин (треугольной нормы дольку затылочной доли) от язычковой извилины и затылочно-височной извилины.
Функция затылочной доли связана с восприятием и переработкой зрительной информации, организацией сложных процессов зрительного восприятия – При этом в области клина проецируется верхняя половина сетчатки глаза, воспринимающая свет от нижних полей зрения; в области язычковой извилины находится нижняя половина сетчатки глаза, воспринимающая свет от верхних полей зрения.
В затылочной коре расположена первичная зрительная область (кора части клиновидной извилины и язычковой дольки, поле 17). Здесь имеется топическое представительство рецепторов сетчатки. Каждой точке сетчатки соответствует свой участок зрительной коры, при этом зона желтого пятна имеет сравнительно большую зону представительства. В связи с неполным перекрестом зрительных путей в зрительную область каждого полушария проецируются одноименные половины сетчатки. Наличие в каждом полушарии проекции сетчатки обоих глаз является основой бинокулярного зрения. Около поля 17 расположена кора вторичной зрительной области (поля 18 и 19). Нейроны этих зон полимодальны и отвечают не только на световые, но и на тактильные и слуховые раздражители. В данной зрительной области происходит синтез различных видов чувствительности, возникают более сложные зрительные образы и осуществляется их распознавание.
Островок, лимбическая кора
Островок, или так называемая закрытая долька, находится в глубине боковой борозды. От примыкающих соседних отделов островок отделен круговой бороздой. Поверхность островка разделена его продольной центральной бороздой на переднюю и заднюю части. В островке проецируется анализатор вкуса.
Лимбическая кора. На внутренней поверхности полушарий над мозолистым телом находится поясная извилина. Эта извилина перешейком позади мозолистого тела переходит в извилину около морского конька – парагиппокампову извилину. Поясная извилина вместе с парагиппокамповой извилиной составляют сводчатую извилину.
Внутренняя и нижняя поверхности полушарий объединяются в так называемую лимбическую (краевую) кору вместе с миндалевидным ядром из группы подкорковых ядер, обонятельным трактом и луковицей, участками лобных, височных и теменных долей коры больших полушарий, а также с подбугровой областью и ретикулярной формацией ствола. Лимбическая кора объединяется в единую функциональную систему – лимбико-ретикулярный комплекс. Основной функцией этих отделов мозга является не столько обеспечение связи с внешним миром, сколько регуляция тонуса коры, влечений и аффективной жизни. Они регулируют сложные, многоплановые функции внутренних органов и поведенческие реакции. Лимбико-ретикулярный комплекс – важнейшая интегративная система организма. Лимбическая система имеет также важное значение в формировании мотиваций. Мотивация (или внутреннее побуждение) включает в себя сложнейшие инстинктивные и эмоциональные реакции (пищевые, оборонительные, половые). Лимбическая система принимает участие также в регуляции сна и бодрствования.
Лимбическая кора выполняет также важную функцию обоняния. Обоняние – восприятие находящихся в воздухе химических веществ. Обонятельный мозг человека обеспечивает обоняние, а также организацию сложных форм эмоциональных и поведенческих реакций. Обонятельный мозг является частью лимбической системы.
Обонятельный мозг состоит из двух отделов – периферического и центрального. Периферический отдел представлен обонятельным нервом, обонятельными луковицами, первичными обонятельными центрами. Центральный отдел включает извилину морского коня – гиппокамп, зубчатую и сводчатую извилины.
Рецепторный аппарат обоняния расположен в слизистой оболочке носа. По системе нервных проводников информация с рецепторов передается в корковый отдел обонятельного анализатора.
Корковый отдел обонятельного анализатора находится в поясной извилине, извилине морского коня и в крючке морского коня, которые вместе составляют замкнутую кольцевидную область. Периферический отдел обонятельного анализатора связан с корковыми областями обоих полушарий.
Физиологический механизм восприятия запахов обонятельным анализатором окончательно не ясен. Существуют две основные гипотезы, с разных позиций объясняющие природу этого процесса. Согласно одной из гипотез, взаимодействие между молекулами пахучего вещества и хеморецепторами происходит по типу ключа и замка, т.е. типу молекулы соответствует специальный рецептор. Другая гипотеза базируется на предположении о том, что молекулы пахучего вещества имеют определенную волну колебания, на которую «настроены» обонятельные рецепторы. Молекулы, имеющие сходные колебания, должны иметь общую волну и соответственно давать близкие запахи.
Термин «обонятельный мозг» применительно к физиологии человека несколько условен и не раскрывает полностью его многогранной и универсальной функции. «Размещение» центрального звена обонятельного мозга в больших полушариях не случайно и является результатом той огромной «информационной» роли, которую играло обоняние в процессе эволюции при адаптации к внешней среде и регуляции сложных поведенческих реакций. Добывание пищи, выбор особи противоположного пола, забота о потомстве, целостности территории, организации групповых сообществ внутри вида – все эти повседневные функции у многих животных выполняются при непосредственном участии тонко сконструированной системы обонятельной рецепции и основанной на этом способности ряда животных посылать во внешнюю среду тонкие дифференцированные специфические пахучие вещества – сигналы-информаторы.
В жизни людей обоняние потеряло то биологическое информационное значение, которое оно имело у животных. Обонятельная система человека предназначена как для выполнения узкой, «своей», функции, так и для своего рода «зарядки» эмоций. О силе воздействия запахов на эмоциональную сферу, о том, что они являются важнейшим «пищевым субстратом эмоций», хорошо известно с давних времен истории человечества.
Острота обоняния человека может варьировать. Как правило, эти вариации незначительны, однако в отдельных случаях острота обоняния может быть очень высокой (дегустаторы парфюмерной промышленности).
Поскольку обонятельный анализатор играет важную роль в регуляции эмоций, его центральный отдел относят к лимбической системе, образно названной «общим знаменателем» для множества эмоциональных и висцеросоматических реакций организма.
Центр вкусового анализатора располагается в ближайшем соседстве с центром обонятельного анализатора, т.е. в крючке и аммоновом роге, но, кроме того, в самом нижнем отделе задней центральной извилины (поле 43), а также в островке. Как и обонятельный анализатор, центр обеспечивает проекционную функцию, хранение и распознавание вкусовых образов.
На границе височной, затылочной и теменной долей располагается центр анализатора письменной речи (поле 39), который тесно связан с центром Вернике височной доли, с центром зрительного анализатора затылочной доли, а также с центрами теменной доли. Центр чтения обеспечивает распознавание и хранение образов письменной речи.
Основная часть информации об окружающей среде и внутренней среде организма, поступившая в сенсорную кору, передается для дальнейшей ее обработки в ассоциативную кору, после чего инициируется (в случае необходимости) поведенческая реакция с обязательным участием двигательной коры.
Мозолистое тело – дугообразная тонкая пластинка, филогенетически молодая, соединяет срединные поверхности обоих полушарий. Удлиненная средняя часть мозолистого тела сзади переходит в утолщение, а спереди искривляется и дугообразно загибается вниз. Мозолистое тело соединяет филогенетически наиболее молодые участки полушарий и играет важную роль в обмене информацией между ними.
Архитектоника коры больших полушарий мозга
Учение о структурных особенностях строения коры называется архитектоникой. Клетки коры больших полушарий менее специализированы, чем нейроны других отделов мозга; тем не менее определенные их группы анатомически и физиологически тесно связаны с теми или иными специализированными отделами мозга.
Микроскопическое строение
коры головного мозга неодинаково в разных ее отделах. Эти морфологические
различия коры позволили выделить отдельные корковые цитоархитектонические поля.
Имеется несколько вариантов классификаций корковых полей. Большинство
исследователей выделяет 50 цитоархитектонических полей, Микроскопическое
строение их довольно сложное.
Кора состоит из 6 слоев клеток и их волокон. Основной тип строения коры шестислойной, однако, он не везде однороден. Существуют участки коры, где один из слоев выражен значительно, а другой – слабо. В других областях коры намечается подразделение некоторых слоев на подслои и т.д.
Установлено, что области коры, связанные с определенной функцией, имеют сходное строение. Участки коры, которые близки у животных и человека по своему функциональному значению имеют определенное сходство в строении. Те участки мозга, которые выполняют чисто человеческие функции (речь), имеются только в коре человека, а у животных, даже у обезьян, отсутствуют.
Морфологическая и функциональная неоднородность коры головного мозга позволила выделить центры зрения, слуха, обоняния и т.д., которые имеют свою определенную локализацию. Однако неверно говорить о корковом центре как о строго ограниченной группе нейронов. Специализация участков коры формируется в процессе жизнедеятельности. В раннем детском возрасте функциональные зоны коры перекрывают друг друга, поэтому их границы расплывчаты и нечетки. Только в процессе обучения, накопления собственного опыта практической деятельности происходит постепенная концентрация функциональных зон в отделенные друг от друга центры.
Белое вещество больших полушарий состоит из нервных проводников. В соответствии с анатомическими и функциональными особенностями волокна белого вещества делят на ассоциативные, комиссуральные и проекционные.

а – ассоциативные волокна; б – комиссуральные волокна; в-проекционные волокна.
Ассоциативные волокна объединяют различные участки коры внутри одного полушария. Эти волокна бывают короткие и длинные. Короткие волокна обычно имеют дугообразную форму и соединяют соседние извилины. Длинные волокна соединяют отдаленные участки коры. Комиссуальными принято называть те волокна, которые соединяют топографически идентичные участки правого и левого полушарий. Комиссуральные волокна образуют три спайки: переднюю белую спайку, спайку свода, мозолистое тело. Передняя белая спайка соединяет обонятельные области правого и левого полушарий. Спайка свода соединяет между собой гиппокамповые извилины правого и левого полушарий. Основная же масса коммисуальных волокон проходит через мозолистое тело, соединяя между собой симметричные участки обоих полушарий головного мозга.
Проекционными называют те волокна, которые связывают полушария головного мозга с нижележащими отделами мозга – стволом и спинным мозгом. В составе проекционных волокон проходят проводящие пути, несущие афферентную (чувствительную) и эфферентную (двигательную) информацию.
1 – прецентральная извилина; 2 – прецентральная борозда; 3 – верхняя лобная извилина; 4 – центральная борозда; 5 – средняя лобная извилина;
б – нижняя лобная извилина; 7 – восходящая ветвь латеральной борозды;
8 – горизонтальная ветвь латеральной борозды; 9 – задняя ветвь латеральной борозды; 10 – верхняя височная извилина; 11 – средняя височная извилина;
12 – нижняя височная извилина; 13 – теменная долька; 14 – пост-центральная борозда; 15 – постцентральная извилина; 16 – надкраевая извилина;
17 – угловая извилина; 18 – затылочная доля; 19 – мозжечок; 20 – горизонтальная щель мозжечка; 21 – продолговатый мозг
14.1. ОБЩИЕ ПОЛОЖЕНИЯ
Конечный мозг (telencephalon), или большой мозг (cerebrum), располагается в супратенториальном пространстве полости черепа и состоит из двух больших
полушарий (gemispherium cerebralis), разделенных между собой глубокой продольной щелью (fissura longitudinalis cerebri), в которую погружен серп большого мозга (falx cerebri), представляющий собою дубликатуру твердой мозговой оболочки. Большие полушария головного мозга составляют 78% его массы. Каждое из больших полушарий имеет доли: лобную, теменную, височную, затылочную и лимбическую. Они прикрывают структуры промежуточного мозга и находящиеся ниже мозжечкового намета (субтенториально) мозговой ствол и мозжечок.
Каждое из полушарий большого мозга имеет три поверхности: верхнебоковую, или конвекситальную (рис. 14.1а), - выпуклую, обращенную к костям свода черепа; внутреннюю (рис. 14.1б), прилежащую к большому серповидному отростку, и нижнюю, или базальную (рис. 14.1в), повторяющую рельеф основания черепа (передней и средней его ямок) и намета мозжечка. В каждом полушарии различают три края: верхний, нижневнутренний и нижненаружный, и три полюса: передний (лобный), задний (затылочный) и боковой (височный).
Полость каждого большого полушария составляет боковой желудочек мозга, при этом левый боковой желудочек признается первым, правый - вторым. Боковой желудочек имеет центральную часть, находящуюся в глубине теменной доли (lobus parietalis) и три отходящих от нее рога: передний рог проникает в лобную долю (lobus frontalis), нижний - в височную (lobus temporalis), задний - в затылочную (lobus occipitalis). Каждый из боковых желудочков сообщатся с третьим желудочком мозга через межжелудочковое отверстие Монро.
Центральные отделы медиальной поверхности обоих полушарий соединены между собой мозговыми спайками, наиболее массивной из которых является мозолистое тело, и структурами промежуточного мозга.
Конечный мозг, как и другие отделы мозга, состоит из серого и белого вещества. Серое вещество располагается в глубине каждого полушария, формируя там подкорковые узлы, и по периферии свободных поверхностей полушария, где оно составляет кору мозга.
Основные вопросы, связанные со строением, функциями подкорковых узлов и варианты клинической картины при их поражении рассмотрены в главах 5, 6. Кора полушарий большого мозга по площади приблизительно в
Рис. 14.1. Полушария большого мозга.
а - верхнелатеральная поверхность левого полушария: 1 - центральная борозда; 2 - глазничная часть нижней лобной извилины; I - лобная доля; 3 - прецентральная извилина; 4 - прецентральная борозда; 5 - верхняя лобная извилина; 6 - средняя лобная извилина; 7 - покрышечная часть нижней лобной извилины; 8 - нижняя лобная извилина; 9 - латеральная борозда; II - теменная доля: 10 - постцентральная извилина; 11 - постцентральная борозда; 12 - внутритеменная борозда; 13 - надкраевая извилина; 14 - угловая извилина; III - височная доля: 15 - верхняя височная извилина; 16 - верхняя височная борозда; 17 - средняя височная извилина; 18 - средняя височная борозда; 19 - нижняя височная извилина; IV - затылочная доля: б - медиальная поверхность правого полушария: 1 - парацентральная долька, 2 - предклинье; 3 - теменно-затылочная борозда; 4 - клин, 5 - язычная извилина; 6 - латеральная затылочно-височная извилина; 7 - парагиппокампальная извилина; 8 - крючок; 9 - свод; 10 - мозолистое тело; 11 - верхняя лобная извилина; 12 - поясная извилина; в - нижняя поверхность больших полушарий: 1 - продольная межполушарная щель; 2 - глазничные борозды; 3 - обонятельный нерв; 4 - зрительный перекрест; 5 - средняя височная борозда; 6 - крючок; 7 - нижняя височная извилина; 8 - сосцевидное тело; 9 - основание ножки мозга; 10 - латеральная затылочно-височная извилина; 11 - парагиппокампальная извилина; 12 - коллатеральная борозда; 13 - поясная извилина; 14 - язычная извилина; 15 - обонятельная борозда; 16 - прямая извилина.
3 раза превышает видимую при наружном осмотре поверхность полушарий. Это обусловлено тем, что поверхность больших полушарий складчата, имеет многочисленные углубления - борозды (sulci cerebri) и расположенные между ними извилины (gyri cerebri). Кора больших полушарий покрывает всю поверхность извилин и борозд (отсюда другое ее название pallium - плащ), проникая при этом подчас на большую глубину в вещество мозга.
Выраженность и расположение борозд и извилин больших полушарий в определенной степени вариабельны, однако основные из них формируются в процессе онтогенеза и являются постоянными, свойственными каждому нормально развившемуся головному мозгу.
14.2. ОСНОВНЫЕ БОРОЗДЫ И ИЗВИЛИНЫ ПОЛУШАРИЙ БОЛЬШОГО МОЗГА
Верхнебоковая (конвекситальная) поверхность полушарий (рис. 14.1а). Самая большая и глубокая - латеральная борозда (sulcus lateralis), или сильвиева борозда, - отделяет лобную и передние отделы теменной доли от расположенной ниже височной доли. Лобная и теменная доли разграничиваются центральной, или роландовой, бороздой (sulcus centralis), которая прорезает верхний край полушария и направляется по конвекситальной его поверхности вниз и вперед, немного не доходя до латеральной борозды. Теменную долю отделяют от расположенной позади нее затылочной доли проходящие по медиальной поверхности полушария теменно-затылочная и поперечная затылочная борозды.
В лобной доле впереди центральной извилины и параллельно ей расположена предцентральная (gyrus precentralis), или передняя центральная, извилина, которую спереди ограничивает предцентральная борозда (sulcus precentralis). От предцентральной борозды кпереди отходят верхняя и нижняя лобные борозды, разделяющие конвекситальную поверхность передних отделов лобной доли на три лобные извилины - верхнюю, среднюю и нижнюю (gyri frontales superior, media et inferior).
Передний отдел конвекситальной поверхности теменной доли составляет находящаяся позади центральной борозды постцентральная (gyrus postcentralis), или задняя центральная, извилина. Сзади ее окаймляет постцентральная борозда, от которой назад тянется внутритеменная борозда (sulcus intraparietalis), разграничивающая верхнюю и нижнюю теменные дольки (lobuli parietales superior et inferior). В нижней теменной дольке в свою очередь выделяются надкраевая извилина (gyrus supramarginalis), окружающая задний отдел латеральной (сильвиевой) борозды, и угловая извилина (girus angularis), окаймляющая заднюю часть верхней височной извилины.
На конвекситальной поверхности затылочной доли мозга борозды неглубокие и могут существенно варьировать, вследствие этого вариабелен и характер расположенных между ними извилин.
Конвекситальную поверхность височной доли разделяют имеющие почти параллельное с латеральной (сильвиевой) бороздой направление верхняя и нижняя височные борозды, разделяющие конвекситальную поверхность височной доли на верхнюю, среднюю и нижнюю височные извилины (gyri temporales superior, media et inferior). Верхняя височная извилина составляет нижнюю губу латеральной (сильвиевой) борозды. На ее поверхности, обращенной в
сторону боковой борозды, имеется несколько поперечных мелких борозд, выделяющих на ней небольшие поперечные извилины (извилины Гешля), которые можно рассмотреть, лишь раздвинув края латеральной борозды.
Передняя часть латеральной (сильвиевой) борозды представляет собой углубление с широким дном, образующим так называемый островок (insula), или островковую долю (lubus insularis). Прикрьгающий этот островок верхний край боковой борозды называется покрышкой (operculum).
Внутренняя (медиальная) поверхность полушария (рис. 14.1б). Центральная часть внутренней поверхности полушария тесно связана со структурами про- межуточного мозга, от которых ее отграничивают относящиеся к большому мозгу свод (fornix) и мозолистое тело (corpus callosum). Последнее снаружи окаймлено бороздой мозолистого тела (sulcus corporis callosi), начинающейся у передней его части - клюва (rostrum) и заканчивающейся у его утолщенного заднего конца (splenium). Здесь борозда мозолистого тела переходит в глубокую гиппокампову борозду (sulcus hippocampi), которая проникает глубоко в вещество полушария, вдавливая его в полость нижнего рога бокового желудочка, в результате чего и образуется так называемый аммониев рог.
Несколько отступив от борозды мозолистого тела и гиппокамповой борозды, располагаются мозолисто-краевая, подтеменная и носовая борозды, являющиеся продолжением друг друга. Эти борозды отграничивают снаружи дугообразную часть медиальной поверхности полушария мозга, известную как лимбическая доля (lobus limbicus). В лимбической доле различают две извилины. Верхнюю часть лимбической доли составляет верхняя лимбическая (верхняя краевая), или опоясывающая, извилина (girus cinguli), нижняя часть образована нижней лимбической извилиной, или извилиной морского конька (girus hippocampi), или парагиппокампальной извилиной (girus parahyppocampalis), в передней части которой имеется крючок (uncus).
Вокруг лимбической доли мозга расположены образования внутренней поверхности лобной, теменной, затылочной и височной долей. Большую часть внутренней поверхности лобной доли занимает медиальная сторона верхней лобной извилины. На границе между лобной и теменной долями большого полушария расположена парацентральная долька (lobulis paracentralis), которая является как бы продолжением передней и задней центральных извилин на медиальной поверхности полушария. На границе между теменной и затылочной долями хорошо видна теменно-затылочная борозда (sulcus parietooccipitalis). От нижней части ее назад отходит шпорная борозда (sulcus calcarinus). Между этими глубокими бороздами расположена извилина треугольной формы, известная под названием клин (cuneus). Впереди клина находится четырехугольная извилина, относящаяся к теменной доле мозга, - предклинье (precuneus).
Нижняя поверхность полушария (рис. 14.1в). Нижняя поверхность большого полушария состоит из образований лобной, височной и затылочной долей. Прилежащий к средней линии отдел лобной доли составляет прямая извилина (girus rectus). Снаружи ее отграничивает обонятельная борозда (sulcus olphactorius), к которой снизу прилежат образования обонятельного анализатора: обонятельная луковица и обонятельный тракт. Латеральнее его, вплоть до латеральной (сильвиевой) борозды, выходящей на нижнюю поверхность лобной доли, расположены мелкие глазничные извилины (gyri orbitalis). Латеральные отделы нижней поверхности полушария позади латеральной борозды занимает нижняя височная извилина. Медиальнее от нее находится латеральная височно-затылочная извилина (gyrus occipitotemporalis lateralis), или веретенообразная борозда. Перед-
ние отделы ее с внутренней стороны граничат с извилиной гиппокампа, а задние - с язычной (gyrus lingualis) или медиальной височно-затылочной извилиной (gyrus occipitotemporalis medialis). Последняя своим задним концом прилежит к шпорной борозде. Передние отделы веретенообразной и язычной извилин относятся к височной доле, а задние - к затылочной доле мозга.
14.3. БЕЛОЕ ВЕЩЕСТВО БОЛЬШИХ ПОЛУШАРИЙ
Белое вещество больших полушарий состоит из нервных волокон, преимущественно миелиновых, составляющих проводящие пути, обеспечивающие связи между нейронами коры и скоплениями нейронов, образующих таламусы, подкорковые узлы, ядра. Основную часть белого вещества больших полушарий составляет расположенный в его глубине полуовальный центр, или лучистый венец (corona radiata), состоящий главным образом из афферентных и эфферентных проекционных проводящих путей, связующих кору больших полушарий с подкорковыми узлами, ядрами и ретикулярной субстанцией промежуточного мозга и мозгового ствола, с сегментами спинного мозга. Особенно компактно они располагаются между таламусом и подкорковыми узлами, где формируют описанную в главе 3 внутреннюю капсулу.
Нервные волокна, соединяющие между собой участки коры одного полушария, называются ассоциативными. Чем короче эти волокна и образуемые ими связи, тем поверхностнее они располагаются; более длинные ассоциативные связи, расположенные глубже, соединяют между собой относительно отдаленные участки коры большого полушария (рис. 14.2 и 14.3).
Волокна, соединяющие между собой полушария большого мозга и имеющие в связи с этим общую поперечную направленность, называются комиссуральными, или спаечными. Комиссуральные волокна соединяют идентичные участки больших полушарий, создавая возможность сочетанности их функций. Они образуют три спайки большого мозга: самая массивная из них - мозолистое тело (corpus callosum), кроме того, комиссуральные волокна составляют переднюю спайку, расположенную под клювом мозолистого тела (rostrum corporis collosum) и соединяющую обе обонятельные области, а также спайку свода (commissura fornicis), или гиппокамповую спайку, образованную волокнами, соединяющими между собой структуры аммоновых рогов обоих полушарий.
В переднем отделе мозолистого тела проходят волокна, соединяющие между собой лобные доли, далее - волокна, соединяющие теменные и височные доли, задний отдел мозолистого тела связывает между собой затылочные доли мозга. Передняя спайка и спайка свода объединяют главным образом участки древней и старой коры обоих полушарий, передняя спайка, кроме того, обеспечивает связь между их средними и нижними височными извилинами.
14.4. ОБОНЯТЕЛЬНАЯ СИСТЕМА
В процессе филогенеза с развитием большого мозга сопряжено формирование обонятельной системы, функции которой способствуют сохранению жизнеспособности животных и имеют немаловажное значение для жизнедеятельности человека.
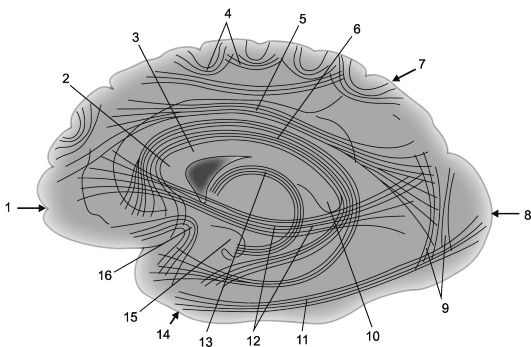
Рис. 14.2. Ассоциационные корково-корковые связи в больших полушариях [по В.П. Воробьеву].
1 - лобная доля; 2 - колено мозолистого тела; 3 - мозолистое тело; 4 - дугообразные волокна; 5 - верхний продольный пучок; 6 - поясная извилина; 7 - теменная доля, 8 - затылочная доля; 9 - вертикальные пучки Вернике; 10 - валик мозолистого тела;
11 - нижний продольный пучок; 12 - подмозолистый пучок (лобно-затылочный нижний пучок); 13 - свод; 14 - височная доля; 15 - крючок извилины гиппокампа; 16 - пучки крючка (fasciculus uncinatus).
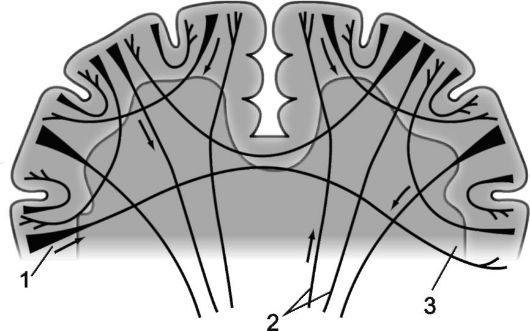
Рис. 14.3. Миелоархитектоника больших полушарий.
1 - проекционные волокна; 2 - комиссуральные волокна; 3 - ассоциативные волокна.
14.4.1. Строение обонятельной системы
Тела первых нейронов обонятельной системы расположены в слизистой оболочке носа, главным образом верхней части носовой перегородки и верхнего носового хода. Обонятельные клетки биполярны. Дендриты их выходят на поверхность слизистой оболочки и заканчиваются здесь специфическими рецепторами, а аксоны группируются в так называемые обонятельные нити (fili olfactorii), число которых с каждой стороны около двадцати. Такой пучок обонятельных нитей и составляет I черепной, или обонятельный, нерв (рис. 14.4). Эти нити проходят в переднюю (обонятельную, ольфакторную) черепную ямку через решетчатую кость и заканчиваются у клеток расположенных здесь обонятельных луко- виц. Обонятельные луковицы и проксимальнее расположенные обонятельные тракты являются, по сути, следствием формирующихся в процессе онтогенеза выпячиваний вещества большого мозга и представляют собой относящиеся к нему структуры.
В обонятельных луковицах находятся клетки, являющиеся телами вторых нейронов обонятельного пути, аксоны которых формируют обонятельные тракты (tracti olfactorii), расположенные под обонятельными бороздками, латеральнее прямых извилин, находящихся на базальной поверхности лобных долей. Обонятельные тракты направляются назад к подкорковым обонятельным центрам. Подходя к передней продырявленной пластинке, волокна обонятельного тракта разделяются на медиальные и латеральные пучки, образующие с каждой стороны по обонятельному треугольнику. В дальнейшем эти волокна подходят к телам третьих нейронов обонятельного анализатора, находящимся
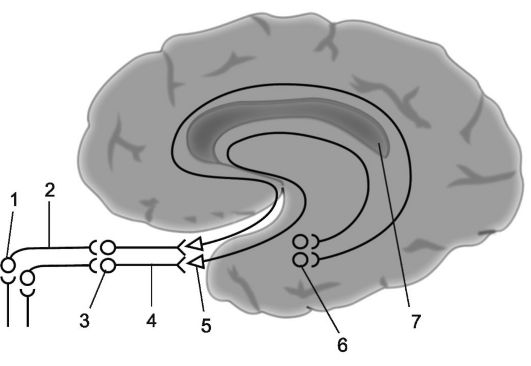
Рис. 14.4. Обонятельный анализатор.
1 - обонятельные клетки; 2 - обонятельные нити (в сумме они составляют обонятельные нервы); 3 - обонятельные луковицы; 4 - обонятельные тракты; 5 - обонятельные треугольники; 6 - парагиппокамповая извилина; 7 - проекционная зона обонятельного анализатора (упрощенная схема).
в периминдалевидной и подмозолистой областях, в ядрах прозрачной перегородки, расположенной кпереди от передней спайки. Передняя спайка соединяет обе обонятельные области, а также обеспечивает их связь с лимбической системой мозга. Часть аксонов третьих нейронов обонятельного анализатора, проходя через переднюю спайку мозга, совершает перекрест.
Аксоны третьих нейронов обонятельного анализатора, расположенных в подкорковых обонятельных центрах, направляются к старой в филогенетическом отношении коре медиобазальной поверхности височной доли (к грушевидной и парагиппокампальной извилинам и к крючку), где расположена проекционная обонятельная зона, или корковый конец обонятельного анализатора (поле 28, по Бродману).
Система обоняния, таким образом, является единственной системой чувствительности, в которой специфические импульсы на пути от рецепторов к коре минуют таламус. Вместе с тем обонятельная система имеет особенно выраженные связи с лимбическими структурами мозга, а получаемая благодаря ей информация оказывает значительное воздействие на состояние эмоциональной сферы и функции вегетативной нервной системы. Запахи могут быть приятными и неприятными, они влияют на аппетит, настроение, могут вызывать разнообразные вегетативные реакции, в частности тошноту, рвоту.
14.4.2. Исследование обоняния и значение его нарушений для топической диагностики
При исследовании состояния обоняния надо выяснить, ощущает ли больной запахи, одинаковы ли эти ощущения с обеих сторон, дифференцирует ли больной характер ощущаемых запахов, нет ли у него обонятельных галлюцинаций - приступообразно возникающих ощущений запаха, отсутствующего в окружающей среде.
Для исследования обоняния пользуются пахучими веществами, запах которых не является резким (резкие запахи могут обусловить раздражение расположенных в слизистой оболочке носа рецепторов тройничного нерва) и заведомо известен больному (в противном случае трудно распознать извращение обоняния). Обоняние проверяется с каждой стороны в отдельности, другую ноздрю при этом необходимо закрыть. Можно пользоваться специально при- готовленными наборами слабых растворов пахучих веществ (мяты, дегтя, камфоры и т.п.), в практической работе могут быть использованы и подручные средства (ржаной хлеб, мыло, банан и т.п.).
Понижение обоняния - гипосмия, отсутствие обоняния - аносмия, обостренное обоняние - гиперосмия, извращение запахов - дизосмия, ощущение запаха при отсутствии стимула - паросмия, субъективное ощущение неприятного запаха, имеющегося на самом деле и обусловленного органической патологией в носоглотке - какосмия, не существующие реально запахи, которые пациент ощущает пароксизмально - обонятельные галлюцинации, - чаще являются обонятельной аурой височной эпилепсии, которая может быть обусловлена разными причинами, в частности опухолью височной доли.
Гипосмия или аносмия с обеих сторон обычно является следствием поражения слизистой оболочки носа, обусловленного острым катаральным состоянием, гриппом, аллергическим ринитом, атрофией слизистой оболочки
носа вследствие хронического ринита и длительного применения вазоконстрикторных капель в нос. Хронический ринит с атрофией слизистой оболочки носа (атрофический ринит), болезнь Шегрена обрекают человека на стойкую аносмию. Двусторонняя гипосмия может быть обусловлена гипотиреозом, сахарным диабетом, гипогонадизмом, почечной недостаточностью, длительным контактом с тяжелыми металлами, формальдегидом и т.п.
Вместе с тем односторонняя гипосмия или аносмия нередко оказывается следствием внутричерепной опухоли, чаще менингиомы передней черепной (ольфакторной) ямки, которая составляет до 10% внутричерепных менингиом, а также некоторых глиальных опухолей лобной доли. Обонятельные расстройства возникают вследствие сдавления обонятельного тракта на стороне патологического очага и могут в течение определенного времени быть единственным очаговым симптомом заболевания. Визуализацию опухолей может обеспечить КТили МРТ-сканирование. По мере увеличения менингиомы ольфактор- ной ямки, как правило, развиваются психические нарушения, характерные для лобного синдрома (см. главу 15).
Одностороннее поражение отделов обонятельного анализатора, расположенных выше его подкорковых центров, ввиду неполного перекреста проводящих путей на уровне передней мозговой спайки обычно не ведет к значительному снижению обоняния. Раздражение патологическим процессом коры медиобазальных отделов височной доли, прежде всего парагиппокампальной извилины и ее крючка, может обусловить приступообразное возникновение обонятельных галлюцинаций. Больной вдруг начинает ощущать без повода к тому запах, чаще неприятного характера (запах горелого, прелого, гнилого, жженого и т.п.). Обонятельные галлюцинации при наличии в медиобазальных отделах височной доли мозга эпилептогенного фокуса могут быть проявлением ауры эпилептического припадка. Поражение проксимального отдела, в частности коркового конца обонятельного анализатора, может обусловить умеренную двустороннюю (больше на противоположной стороне) гипосмию и нарушение способности к идентификации и дифференцировке запахов (обонятельная агнозия). Последняя форма обонятельного расстройства, проявляющаяся в старческом возрасте, скорее всего связана с нарушением функции коры вследствие атрофических процессов в ее проекционной обонятельной зоне.
14.5. ЛИМБИКО-РЕТИКУЛЯРНЫЙ КОМПЛЕКС
В 1878 г. П. Брока (Broca P., 1824-1880) под названием «большая краевая, или лимбическая, доля» (от лат. limbus - край) объединил гиппокамп и поясную извилину, соединенные между собой посредством перешейка поясной извилины, расположенной над валиком мозолистого тела.
В 1937 г. Д. Папец (Papez J.) на основании экспериментальных данных вы- двинул аргументированное возражение против существовавшей ранее концепции о причастности медиобазальных структур больших полушарий главным образом к обеспечению обоняния. Он предположил, что основная часть медиобазальных отделов большого полушария, называемых тогда обонятельным мозгом (rhinencephalon), к которому относится лимбическая доля, представляет собой морфологическую основу нервного механизма аффективного поведения, и объединил их под названием «эмоциональный круг», в который включил гипоталамус,
передние ядра таламуса, поясную извилину, гиппокамп и их связи. С тех пор эти структуры физиологами стали именоваться также кругом Папеца.
Понятие «висцеральный мозг» предложил P.D. McLean (1949), обозначив таким образом сложное анатомо-физиологическое объединение, которое с 1952 г. стали именовать «лимбической системой». Позже выяснилось, что лимбическая система участвует в выполнении многообразных функций, и сейчас большую ее часть, включающую поясную и гиппокампову (парагиппокампальную) извилины, принято объединять в лимбическую область, которая имеет многочисленные связи со структурами ретикулярной формации, составляя вместе с ней лимбико-ретикулярный комплекс, обеспечивающий широкий круг физиологических и психологических процессов.
В настоящее время к лимбической доле принято относить элементы старой коры (archiocortex), покрывающей зубчатую извилину и гиппокампову извилину; древнюю кору (paleocortex) переднего отдела гиппокампа; а также среднюю, или промежуточную, кору (mesocortex) поясной извилины. Термин «лимбическая система» включает компоненты лимбической доли и связанных с ней структур - энторинальную (занимающую большую часть парагиппокампальной извилины) и септальную области, а также миндалевидный комплекс и сосцевидное тело (Дуус П., 1995).
Сосцевидное тело связывает структуры этой системы со средним мозгом и с ретикулярной формацией. Импульсы, возникающие в лимбической системе, могут передаваться через переднее ядро таламуса в поясную извилину и к новой коре по путям, образованным ассоциативными волокнами. Импульсы, возникающие в гипоталамусе, могут достигать орбитофронтальной коры и медиального дорсального ядра таламуса.
Многочисленные прямые и обратные связи обеспечивают взаимосвязь и взаимозависимость лимбических структур и многих образований промежуточного мозга и оральных отделов ствола (неспецифические ядра таламуса, гипоталамус, скорлупа, уздечка, ретикулярная формация ствола мозга), а также с подкорковыми ядрами (бледный шар, скорлупа, хвостатое ядро) и с новой корой больших полушарий, прежде всего с корой височной и лобной долей.
Несмотря на филогенетические, морфологические и цитоархитектонические различия, многие из упомянутых структур (лимбическая область, центральные и медиальные структуры таламусов, гипоталамус, ретикулярная формация ствола) принято включать в так называемый лимбико-ретикулярный комплекс, который выступает как зона интеграции множества функций, обеспечивая организацию полимодальных, целостных реакций организма на разнообразные воздействия, что особенно ярко проявляется в условиях стрессовых ситуаций.
Структуры лимбико-ретикулярного комплекса имеют большое количество входов и выходов, через которые проходят замкнутые круги из многочисленных афферентных и эфферентных связей, обеспечивающие сочетанное функционирование входящих в этот комплекс образований и взаимодействие их со всеми отделами головного мозга, включая кору больших полушарий.
В структурах лимбико-ретикулярного комплекса происходит конвергенция чувствительной импульсации, возникающей в интеро- и экстерорецепторах, включая рецепторные поля органов чувств. На этой основе в лимбико-ретикулярном комплексе происходит первичный синтез информации о состоянии внутренней среды организма, а также о воздействующих на организм факторах внешней среды, и формируются элементарные потребности, биологические мотивации и сопутствующие им эмоции.
Лимбико-ретикулярный комплекс определяет состояние эмоциональной сферы, участвует в регуляции вегетативно-висцеральных соотношений, направленных на поддержание относительного постоянства внутренней среды (гомеостаза), а также энергетическое обеспечение и корреляцию двигательных актов. От его состояния зависит уровень сознания, возможность автоматизированных движений, активность двигательных и психических функций, речи, внимание, способность к ориентировке, память, смена бодрствования и сна.
Поражение структур лимбико-ретикулярного комплекса может сопровождаться разнообразной клинической симптоматикой: выраженными изменениями в эмоциональной сфере перманентного и пароксизмального характера, анорексией или булимией, сексуальными расстройствами, нарушением памяти, в частности признаками синдрома Корсакова, при котором больной теряет способность к запоминанию текущих событий (текущие события удерживает в памяти не более 2 мин), вегетативно-эндокринными расстройствами, нарушениями сна, психосенсорными расстройствами в виде иллюзий и галлюцинаций, изменениями сознания, клиническими проявлениями акинетического мутизма, эпилептическими припадками.
К настоящему времени проведено большое количество исследований, посвященных изучению морфологии, анатомических связей, функции лимбической области и других структур, включаемых в лимбико-ретикулярный комплекс, однако физиология и особенности клинической картины его поражения и на сегодняшний день во многом нуждаются в уточнении. Большинство данных о его функции, особенно функции парагиппокампальной области, получено в экспериментах на животных методами раздражения, экстирпации или стереотаксиса. Полученные таким образом результаты требуют осторожности при их экстраполяции на человека. Особое значение имеют клинические наблюдения за больными с поражением медиобазальных отделов большого полушария.
В 50-60-х годах XX в. в период развития психохирургии появились сообщения о лечении больных с некурабельными психическими расстройствами и хроническим болевым синдромом путем двусторонней цингулотомии (рассечение поясных извилин), при этом обычно отмечали регресс тревоги, обсессивных состояний, психомоторного возбуждения, болевых синдромов, что признавалось доказательством участия поясной извилины в формировании эмоций и боли. Вместе с тем бицингулотомия вела к глубоким личностным нарушениям, к дезориентации, снижению критичности своего состояния, эйфории.
Анализ 80 верифицированных клинических наблюдений поражения гиппокампа на базе Нейрохирургического института РАМН приведен в монографии Н.Н. Брагиной (1974). Автор приходит к выводу, что височный медиобазальный синдром включает висцеровегетативные, двигательные и психические нарушения, проявляющиеся обычно в комплексе. Все многообразие клинических проявлений Н.Н. Брагина сводит к двум основным многофакторным вариантам патологии с преобладанием «ирритативных» и «тормозных» феноменов.
К первому из них относятся эмоциональные расстройства, сопровождающиеся двигательным беспокойством (повышенная возбудимость, многоречивость, суетливость, ощущение внутренней тревоги), пароксизмами страха, витальной тоски, многообразными висцеровегетативными расстройствами (изменения пульса, дыхания, желудочно-кишечные нарушения, повышение температуры, усиленное потоотделение и т.д.). У этих больных на фоне постоянного двигательного беспокойства нередко возникали приступы моторного возбужде-
ния. ЭЭГ этой группы больных характеризовалось негрубыми общемозговыми изменениями в сторону интеграции (учащенный и заостренный альфа-ритм, диффузные бета-колебания). Повторные афферентные раздражения при этом вызывали четкие ЭЭГ-реакции, которые в отличие от нормальных не угасали по мере повторно предъявляемых раздражителей.
Второй («тормозной») вариант медиобазального синдрома характеризуют эмоциональные нарушения в форме депрессии с двигательной заторможенностью (подавленный фон настроения, обеднение и замедление темпа психических процессов, изменения моторики, напоминающие по типу акинетико-ригидный синдром. Менее характерны отмеченные в первой группе висцеровегетативные пароксизмы. ЭЭГ больных этой группы характеризовалось общемозговыми изменениями, проявляющимися в преобладании медленных форм активности (нерегулярный, замедленный альфа-ритм, группы тета-колебаний, диффузные дельта-волны). Обращало на себя внимание резкое снижение реактивности ЭЭГ.
Между этими двумя крайними вариантами были и промежуточные с переходными и смешанными сочетаниями отдельных симптомов. Так, при некоторых из них характерны относительно слабые признаки ажитированной депрессии с повышенной двигательной активностью и утомляемостью, с превалированием сенестопатических ощущений, подозрительностью, доходящей у некоторых больных до параноидальных состояний, ипохондрического бреда. Другая промежуточная группа отличалась крайней интенсивностью депрессивной симптоматики на фоне скованности больного.
Эти данные позволяют говорить о двояком (активирующем и тормозном) влиянии гиппокампа и других структур лимбической области на поведенческие реакции, эмоции, особенности психического статуса и биоэлектрическую активность коры. В настоящее время сложные клинические синдромы такого типа не следует расценивать как первично-очаговые. Скорее их нужно рассматривать в свете представлений о многоуровневой системе организации моз- говой деятельности.
С.Б. Буклина (1997) привела данные обследования 41 больного с артериовенозными мальформациями в зоне расположения поясной извилины. До операции у 38 больных на первый план выступали расстройства памяти, причем у пятерых из них имелись признаки корсаковского синдрома, у трех больных корсаковский синдром возник после операции, при этом выраженность нарастания дефектов памяти коррелировала со степенью разрушения самой поясной извилины, а также с вовлечением в патологический процесс прилежащих структур мозолистого тела, при этом амнестический синдром не зависел от стороны расположения мальформации и ее локализации по длиннику поясной извилины.
Основными характеристиками выявленных амнестических синдромов были расстройства воспроизведения слухоречевых стимулов, нарушения избирательности следов в виде включений и контаминаций, неудержания смысла при передаче рассказа. У большинства больных была снижена критичность оценки своего состояния. Автором отмечено сходство этих нарушений с ам- нестическими дефектами у больных с лобными поражениями, что может быть объяснено наличием связей поясной извилины с лобной долей.
Более распространенные патологические процессы в лимбической области вызывают выраженные расстройства вегетативно-висцеральных функций.
Мозолистое тело (corpus callosum) - наибольшая комиссура между большими полушариями мозга. Передние ее отделы, в частности колено мозолистого
тела (genu corporis callosi), связывают между собой лобные доли, средние отделы - ствол мозолистого тела (truncus corporis callosi) - обеспечивают связь височных и теменных отделов полушарий, задние отделы, в частности валик мозолистого тела (splenium corporis callosi), соединяют затылочные доли.
Поражения мозолистого тела обычно сопровождаются расстройствами состояния психики больного. Разрушение переднего его отдела ведет к развитию «лобной психики» (аспонтанность, нарушения плана действий, поведения, критики, характерные для лобно-каллезного синдрома - акинезия, амимия, аспонтанность, астазия-абазия, апраксия, хватательные рефлексы, деменция). Разобщение связей между теменными долями ведет к извращению понимания «схемы тела» и появлению апраксии главным образом в левой руке. Разобщение височных долей может проявляться нарушением восприятия внешней среды, потерей правильной ориентации в ней (амнестические расстройства, конфабуляции, синдром уже виденного и т.п.). Патологические очаги в задних отделах мозолистого тела нередко характеризуются признаками зрительной агнозии.
14.6. АРХИТЕКТОНИКА КОРЫ МОЗГА
Строение коры полушарий большого мозга неоднородно. Менее сложны по строению рано возникающие в процессе филогенеза древняя кора (archiocortex) и старая кора (paleocortex), относящиеся в основном к лимбической доле мозга. Большая же часть коры полушарий мозга (95,6%) в связи со своим более поздним с точки зрения филогенеза формированием называется новой корой (neocortex) и имеет гораздо более сложное многослойное строение, но тоже неоднотипное в различных ее зонах.
В связи с тем, что архитектоника коры находится в определенной связи с ее функцией, ее изучению уделено много внимания. Одним из основателей учения о цитоархитектонике коры был В.А. Бец (1834-1894), впервые в 1874 г. описавший большие пирамидные клетки двигательной зоны коры (клетки Беца) и определивший принципы разделения коры больших полушарий мозга на основные области. В дальнейшем большой вклад в развитие учения о строении коры внесли многие исследователи - А. Кэмбелл (A. Cambell), Э. Смит (E. Smith), К. Бродманн (K. Brodmann), Оскар Фогт и Сесилия Фогт (O. Vogt, S. Vogt). Большие заслуги в изучении архитектоники коры принадлежат кол- лективу Института мозга АМН (С.А. Саркисов, Н.И. Филимонов, Е.П. Кононова и др.).
Основной тип строения новой коры (рис. 14.5), с которым сравниваются все его участки, - кора, состоящая из 6 слоев (гомотипическая кора, по Бродману).
I слой - молекулярный, или зональный, самый поверхностный, беден клетками, волокна его имеют направление, в основном параллельное поверхности коры.
II слой - наружный зернистый. Состоит из большого количества густо расположенных мелких зернистых нервных клеток.
III слой - малых и средних пирамид, самый широкий. Состоит из пирамидных клеток, размеры которых неодинаковы, что позволяет в большинстве корковых полей делить этот слой на подслои.
IV слой - внутренний зернистый. Состоит из густо расположенных мелких клеток-зерен круглой и угловатой формы. Этот слой наиболее изменчивый, в
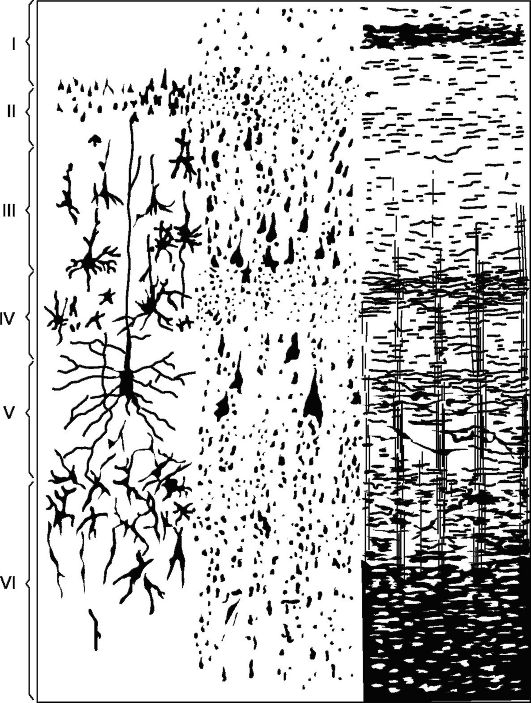
Рис. 14.5. Цитоархитектоника и миелоархитектоника двигательной зоны коры больших полушарий.
Слева: I - молекулярный слой; II - наружный зернистый слой; III - слой малых и средних пирамид; IV - внутренний зернистый слой; V - слой больших пирамид; VI - слой полиморфных клеток; справа - элементы миелоархитектоники.
некоторых полях (например, поле 17) он разделяется на подслои, местами же резко истончается и даже вовсе исчезает.
Vслой - больших пирамид, или ганглионарный. Содержит крупные пирамидные клетки. В некоторых зонах мозга слой делится на подслои, в двигательной зоне он состоит из трех подслоев, средний из которых содержит гигантские пирамидные клетки Беца, достигающие в диаметре 120 мкм.
VIслой - полиморфных клеток, или мультиформный. Состоит, главным образом, из треугольных веретенообразных клеток.
Строение коры мозга имеет большое количество вариаций, обусловленных изменением толщины отдельных слоев, истончением или исчезновением или,
наоборот, утолщением и разделением на подслои некоторых из них (гетеротипические зоны, по Бродману).
Кора каждого большого полушария делится на несколько областей: затылочную, верхнюю и нижнюю теменные, постцентральную, область центральных извилин, предцентральную, лобную, височную, лимбическую, островковую. Каждая из них в соответствии с особенностями подразделяется на ряд полей, причем каждое поле имеет свое условное порядковое обозначение (рис. 14.6).
Изучение архитектоники коры большого мозга наряду с физиологическими, в том числе электрофизиологическими, исследованиями и клиническими наблюдениями во многом содействовало решению вопроса о распределении функций в коре.
14.7. ПРОЕКЦИОННЫЕ И АССОЦИАТИВНЫЕ ПОЛЯ КОРЫ
В процессе развития учения о роли коры больших полушарий и отдельных ее участков в выполнении определенных функций существовали разные, подчас противоположные, точки зрения. Так, существовало мнение о строго локальном представительстве в коре мозга всех свойственных человеку способностей и функций, вплоть до самых сложных, психических (локализационизм, психоморфологизм). Ему противопоставлялось другое мнение об абсолютной функциональной равноценности всех участков коры больших полушарий мозга (эквипотенцализм).
Важный вклад в учение о локализации функций в коре большого мозга внес И.П. Павлов (1848-1936). Он выделял проекционные зоны коры (корковые концы анализаторов отдельных видов чувствительности) и расположенные между ними ассоциативные зоны, изучал в мозге процессы торможения и возбуждения, их влияние на функциональное состояние коры больших полушарий. Деление коры на проекционные и ассоциативные зоны способствует пониманию организации работы коры больших полушарий и оправдывает себя при решении практических задач, в частности при топической диагностике.
Проекционные зоны обеспечивают главным образом простые специфические физиологические акты, прежде всего восприятие ощущений определенной модальности. Подходящие к ним проекционные проводящие пути связывают эти зоны с находящимися с ними в функциональном соответствии рецепторными территориями на периферии. Примерами проекционных корковых зон являются уже описанные в предыдущих главах область задней центральной извилины (зона общих видов чувствительности) или расположенная на медиальной стороне затылочной доли область шпорной борозды (проекционная зрительная зона).
Ассоциативные зоны коры не имеют непосредственных связей с периферией. Они находятся между проекционными зонами и имеют многочисленные ассоциативные связи с этими проекционными и с другими ассоциативными зонами. Функцией ассоциативных зон является осуществление высшего анализа и синтеза многих элементарных и более сложных компонентов. Здесь по существу происходит осмысление поступающей в мозг информации, формирование представлений и понятий.
Г.И. Поляков в 1969 г. на основании сравнения архитектоники коры головного мозга человека и некоторых животных установил, что ассоциативные
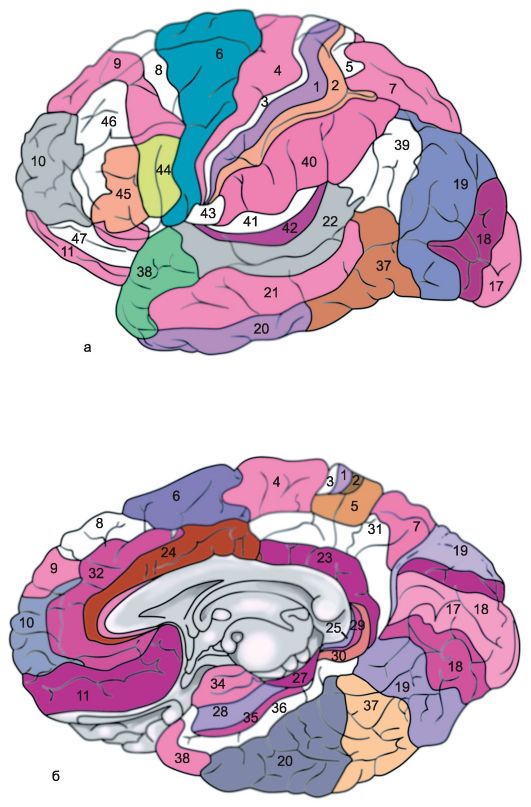
Рис. 14.6. Архитектонические поля коры больших полушарий [по Бродману]. а - наружная поверхность; б - медиальная поверхность.
зоны в коре больших полушарий человека составляют 50%, в коре высших (человекообразных) обезьян - 20%, у низших обезьян этот же показатель равен 10% (рис. 14.7). Среди ассоциативных зон коры человеческого мозга тот же автор предложил выделять вторичные и третичные поля. Вторичные ассоциативные поля примыкают к проекционным. Они осуществляют еще сохраняющий специфическую направленность анализ и синтез элементарных ощущений.
Третичные ассоциативные поля расположены в основном между вторичными и являются зонами перекрытия соседних территорий. Они имеют отношение прежде всего к аналитической деятельности коры, обеспечивая высшие психические функции, свойственные человеку в их наиболее сложных интеллектуальных и речевых проявлениях. Функциональная зрелость третичных ас-
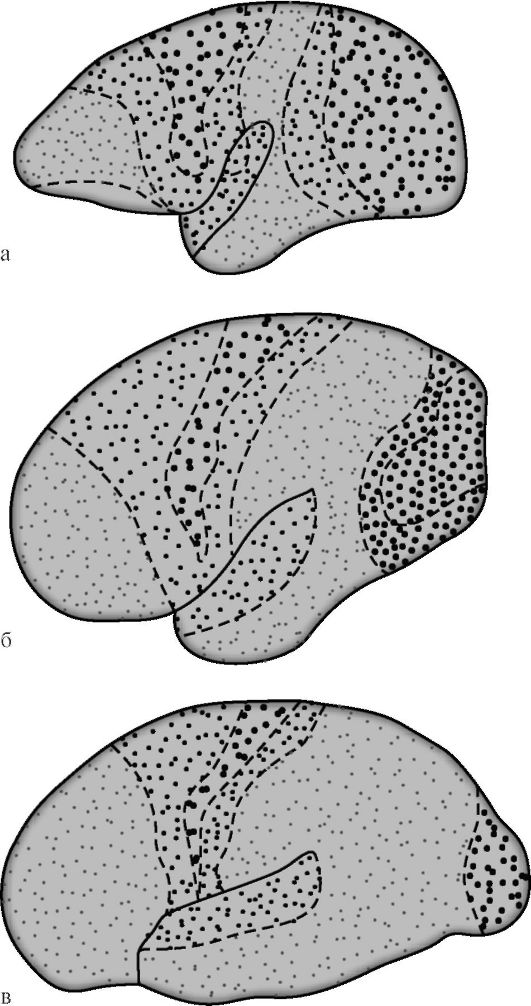
Рис. 14.7. Дифференциация проекционных и ассоциативных зон коры больших полушарий в процессе эволюции приматов [по Г.И. Полякову]. а - мозг низшей обезьяны; б - мозг высшей обезьяны; в - мозг человека. Крупными точками обозначены проекционные зоны, мелкими - ассоциативные. У низших обезьян ассоциативные зоны занимают 10% площади коры, у высших - 20%, у человека - 50%.
социативных полей коры большого мозга наступает наиболее поздно и только в благоприятной социальной среде. В отличие от других корковых полей третичным полям правого и левого полушарий свойственна выраженная функци- ональная асимметрия.
14.8. ТОПИЧЕСКАЯ ДИАГНОСТИКА ПОРАЖЕНИЙ КОРЫ БОЛЬШОГО МОЗГА
14.8.1. Проявления поражения проекционных зон коры больших полушарий
В коре каждого полушария большого мозга позади центральной извилины располагается по 6 проекционных зон.
1. В передней части теменной доли, в области задней центральной извилины (цитоархитектонические поля 1, 2, 3) находится проекционная зона общих видов чувствительности (рис. 14.4). Участки расположенной здесь коры получают чувствительные импульсы, приходящие по проекционным путям общих видов чувствительности, от рецепторного аппарата противоположной половины тела. Чем выше находится участок этой проекционной зоны коры, тем с более низко расположенными частями противоположной половины тела он имеет проекционные связи. Частям тела, имеющим обширную рецепцию (язык, ладонная поверхность кисти), соответствуют неадекватно большие части площади проекционных зон, тогда как другие части тела (проксимальные отделы конечностей, туловище) имеют малую площадь коркового представительства.
Раздражение патологическим процессом корковой зоны общих видов чувствительности ведет к возникновению приступа парестезии в частях тела, соответствующих раздражаемым участкам коры мозга (сенситивный джексоновский припадок), который может развернуться во вторичный генерализованный пароксизм. Поражение коркового конца анализатора общих видов чувствительности может обусловить развитие гипалгезии или анестезии в соответствующей зоне противоположной половины тела, при этом участок гипестезии или анестезии может быть вертикально-циркуляторного или корешково-сегментарного типа. В первом случае расстройство чувствительности проявляется на противоположной относительно патологического очага стороне в области губ, большого пальца или в дистальной части конечности с циркулярной границей, иногда по типу носка или перчатки. Во втором случае зона нарушения чувствительности имеет форму полосы и располагается вдоль внутреннего или наружного края руки или ноги; это объясняется тем, что внутренняя сторона конечностей представлена в передних, а наружная сторона - в задних отделах проекционной зоны анализатора общих видов чувствительности.
2. Зрительная проекционная зона находится в коре медиальной поверхности затылочной доли в области шпорной борозды (поле 17). В этом поле отмечается расслоение IV (внутреннего зернистого) слоя коры пучком миелиновых волокон на два подслоя. Отдельные участки поля 17 получают импульсы от определенных отделов гомонимных половин сетчаток обоих глаз; при этом импульсы, идущие от нижних отделов гомонимных половин сетчаток, достигают
коры нижней губы шпорной борозды, а импульсы, идущие от верхних частей сетчаток, направляются к коре верхней ее губы.
Поражение патологическим процессом зрительной проекционной зоны ведет к возникновению на противоположной стороне квадрантной или полной гомонимной гемианопсии на стороне, противоположной патологическому очагу. Двустороннее поражение корковых полей 17 или идущих к ним проекционных зрительных путей может вести к полной слепоте. Раздражение коры зрительной проекционной зоны может обусловить появление зрительных галлюцинаций в виде фотопсий в соответствующих частях противоположных половин полей зрения.
3. Слуховая проекционная зона расположена в коре извилин Гешля на нижней губе латеральной (сильвиевой) борозды (поля 41 и 42), являющихся, по сути, частью верхней височной извилины. Раздражение этой зоны коры может обусловить возникновение слуховых галлюцинаций (приступы ощущения шума, звона, свиста, жужжания и т.п.). Разрушение слуховой проекционной зоны с одной стороны может обусловить некоторое снижение слуха на оба уха, в большей степени на противоположное относительно патологического очага.
4 и 5. Обонятельная и вкусовая проекционные зоны находятся на медиальной поверхности сводчатой извилины (лимбической области) мозга. Первая из них располагается в парагиппокампальной извилине (поле 28). Проекционную зону вкуса принято локализовать в коре оперкулярной области (поле 43). Раздражение проекционных зон обоняния и вкуса может обусловить их извращение или же ведет к развитию соответствующих обонятельных и вкусовых галлюцинаций. Одностороннее выпадение функции проекционных зон обоняния и вкуса может обусловить некоторое снижение соответственно обоняния и вкуса с обеих сторон. Двустороннее разрушение корковых концов тех же анализаторов проявляется отсутствием с обеих сторон соответственно обоняния и вкуса.
6. Вестибулярная проекционная зона. Локализация ее не уточнена. В то же время известно, что вестибулярный аппарат имеет многочисленные анатомические и функциональные связи. Возможно, что локализация представительства вестибулярной системы в коре до сих пор не уточнена потому, что оно полифокально. Н.С. Благовещенская (1981) считает, что в коре мозга вестибулярные проекционные зоны представлены несколькими анатомо-функциональными, взаимодействующими между собой комплексами, которые располагаются в поле 8, в месте стыка лобной, височной и теменной долей и в зоне центральных извилин, при этом предполагается, что каждый из этих участков коры выполняет свойственные ему функции. Поле 8 - произвольный центр взора, раздражение его вызывает поворот взора в сторону, противоположную патологическому очагу, изменения ритма и характера экспериментального нистагма, особенно вскоре после эпилептического припадка. В коре височной доли располагаются структуры, раздражение которых вызывает головокружение, что проявляется, в частности, при височной эпилепсии; поражение участков представительства вестибулярных структур в коре центральных извилин сказывается на состоянии тонуса поперечнополосатой мускулатуры. Клинические наблюдения позволяют считать, что ядерно-корковые вестибулярные пути совершают частичный перекрест.
Следует подчеркнуть, что признаки раздражения перечисленных проекционных зон могут быть проявлением соответствующей по характеру ауры эпилептического припадка.
И.П. Павлов считал возможным рассматривать кору предцентральной извилины, влияющую на двигательные функции и тонус мышц преимущественно противоположной половины тела, с которыми ее связывают прежде всего корково-ядерные и корково-спинномозговые (пирамидные) проводящие пути, как проекционную зону так называемого двигательного анализатора. Эта зона занимает прежде всего поле 4, на которое в основном и проецируется противоположная половина тела в перевернутом виде. В этом поле содержатся основная масса гигантских пирамидных клеток (клетки Беца), аксоны которых составляют 2-2,5% всех волокон пирамидного пути, а также средние и мелкие пирамидные клетки, которые вместе с аксонами таких же клеток, расположенных в прилежащем к полю 4 более обширном поле 6, участвуют в осуществлении моносинаптических и полисинаптических корково-мышечных связей. Моносинаптические связи обеспечивают главным образом быстрые и точные целенаправленные действия, зависящие от сокращений отдельных поперечно- полосатых мышц.
Поражение нижних отделов моторной зоны обычно ведет к развитию на противоположной стороне брахиофациального (плечелицевого) синдрома или лингвофациобрахиального синдрома, которые часто наблюдаются у больных с нарушением мозгового кровообращения в бассейне средней мозговой артерии, при этом выявляется сочетанный парез мышц лица, языка и руки, прежде всего плеча по центральному типу.
Раздражение коры двигательной зоны (поля 4 и 6) ведет к появлению судорог в проецируемых на эту зону мышцах или мышечных группах. Чаще это локальные судороги по типу джексоновской эпилепсии, которые могут трансформироваться во вторично-генерализованный эпилептический припадок.
14.8.2. Проявления поражения ассоциативных полей коры больших полушарий
Между проекционными зонами коры находятся ассоциативные поля. К ним поступают импульсы главным образом от клеток проекционных зон коры. В ассоциативных полях происходит анализ и синтез информации, подвергшей- ся первичной обработке в проекционных полях. Ассоциативные зоны коры верхней теменной дольки обеспечивают синтез элементарных ощущений, в связи с этим здесь формируются такие сложные виды чувствительности, как чувство локализации, чувство веса, двухмерно-пространственное чувство, а также сложные кинестетические ощущения.
В области межтеменной борозды расположена ассоциативная зона, обеспечивающая синтез ощущений, исходящих из частей собственного тела. Поражение этого участка коры ведет к аутотопагнозии, т.е. к неузнаванию или же игнорированию частей собственного тела, или к псевдомелии - ощущению наличия дополнительной руки или ноги, а также к анозогнозии - отсутствию осознавания физического дефекта, возникшего в связи с болезнью (например, паралича или пареза конечности). Обычно все виды аутотопагнозии и анозогнозия возникают при расположении патологического процесса справа.
Поражение нижней теменной дольки может проявляться расстройством синтеза элементарных ощущений или невозможностью сопоставления синтезированных сложных ощущений с «бывшим когда-то в восприятии подоб-
ным же образом, на основании результатов которого и происходит узнавание» (В.М. Бехтерев). Это проявляется нарушением двухмерно-пространственного чувства (графоестезии) и трехмерно-пространственного чувства (стереогноза) - астереогнозом.
В случае поражения премоторных зон лобной доли (поля 6, 8, 44) обычно возникает лобная атаксия, при которой нарушается синтез афферентной импульсации (кинестетической афферентации), сигнализирующей о меняющемся в процессе совершаемых движений положении частей тела в пространстве.
При нарушении функции коры передних отделов лобной доли, имеющей связи с противоположным полушарием мозжечка (лобно-мосто-мозжечковые связи), на противоположной патологическому очагу стороне возникают статокинетические нарушения (лобная атаксия). Особенно отчетливыми бывают нарушения поздно развивающихся форм статокинетики - прямостояния и прямохождения. В результате у больного возникают неуверенность, шаткость походки. Во время ходьбы корпус его отклоняется назад (симптом Хеннера), ступни он ставит на одной прямой (лисья походка), иногда при ходьбе отмечается «заплетание» ног. У некоторых больных с поражением передних отделов лобных долей развивается своеобразное явление: при отсутствии параличей и парезов и сохранной возможности совершать движения ногами в полном объеме больные не могут стоять (астазия) и ходить (абазия).
Поражение ассоциативных зон коры часто характеризуется развитием клинических проявлений нарушения высших психических функций (см. главу 15).
Большие полушария являются высшим отделом центральной нервной системы. Это самый крупный отдел мозга. Они представляют собой парные образования, объединенные мозолистым телом, которое представляет собой тяж нервных волокон. У взрослого человека большие полушария составляют до 80 % массы головного мозга. Сверху они покрыты серым веществом - корой больших полушарий . В коре насчитывается 12 -18 млрд нервных клеток. Площадь поверхности коры больших полушарий у взрослого человека достигает 2200-2600 см 2 . Рассмотрим строение и значение больших полушарий головного мозга в этой статье.
Это интересно. Верно ли, что, чем больше размер мозга, тем более высокий интеллект у его обладателя? Вне всякого сомнения, чем больше нервных клеток («серого вещества») имеется в мозге, тем более сложными могут быть связи между ними и тем более высоким интеллектом может обладать человек. Но вместе с тем одних только размеров недостаточно. Более важна организация мозга. Например, выдающийся писатель Иван Тургенев имел мозг массой 1700 г, а не менее выдающийся Анатоль Франс - всего 1100 г. При этом средняя масса мозга человека 1400-1500 г.
Практически все навыки, приобретаемые человеком в течение жизни, так или иначе связаны с функциями коры больших полушарий. Кора является материальной основой психики. Она обеспечивает речевую, мыслительную деятельность и память.
Многочисленные борозды (углубления) делят полушария на выпуклые извилины (складки) и доли. Складчатое строение существенно увеличивает площадь поверхности и объем коры.
Три главные борозды - центральная , боковая и теменно-затылочная - разграничивают каждое полушарие головного мозга на четыре доли: лобную, теменную, затылочную и височную (рис.). Доли, в свою очередь, расчленяются бороздами на множество извилин.
Рис. Кора больших полушарий
Разные участки коры больших полушарий выполняют различные функции, поэтому их делят на зоны. Различают сенсорные (чувствительные), ассоциативные и двигательные (моторные) зоны.
Сенсорные зоны являются высшими центрами различных видов чувствительности. При их раздражении возникают ощущения, а при поражении наступает нарушение сенсорных функций (слепота, глухота и др.).
В затылочной области коры находится зрительная, в височной - обонятельная, вкусовая и слуховая сенсорные зоны (рис.).
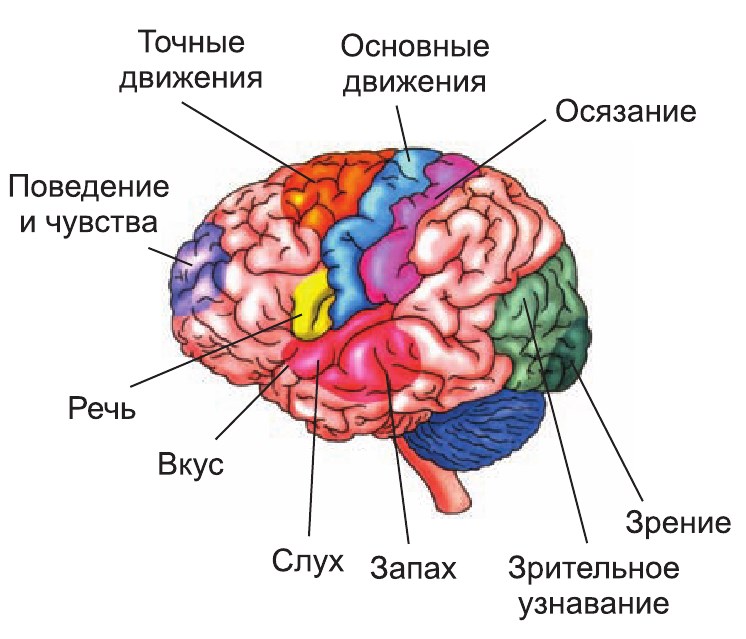
Рис. Локализация функций в коре больших полушарий
Зоны кожного и мышечного чувств располагаются за центральной бороздой, а двигательная зона - перед ней. Самые большие размеры имеют сенсорные зоны кистей рук и лица. И это понятно, учитывая значение этих органов в жизни человека. Наименьшие размеры у сенсорных зон туловища, бедра и голени.
При поступлении импульсов в сенсорные зоны возбуждение возникает также и в ассоциативных . Более того, в одну и ту же ассоциативную зону возбуждение может приходить от разных органов чувств. Так, например, в зрительной ассоциативной зоне возбуждение появляется не только в ответ на зрительные, но и на слуховые раздражения. При нарушении функций ассоциативных зон человек утрачивает способность правильно оценивать явления и события.
Особенно важную роль в сложных формах поведения играют лобные ассоциативные зоны коры. Они обеспечивают обработку сенсорной информации и формируют цель и программу действий. Программа состоит из команд, которые направляются к исполнительным органам. От них информация возвращается в лобные ассоциативные зоны, где определяется, достигнута цель или нет. В последнем случае команда корректируется. С развитием этих долей коры в значительной мере связан высокий уровень психических способностей человека по сравнению с животными.
Двигательными зонами называются отделы коры больших полушарий, которые осуществляют управление произвольными движениями. Двигательная функция различных частей тела представлена в передней центральной извилине. Наибольшее пространство в ней занимают двигательные зоны кистей, пальцев рук и мышц лица, а наименьшее - мышц туловища.
Функции полушарий
Правое и левое полушария у человека выполняют разные функции. В левом полушарии находятся центры устной и письменной речи. Здесь же осуществляются процессы анализа и синтеза информации, делаются обобщения и принимаются решения. Обеспечиваемое левым полушарием словесно-логическое мышление позволяет познать сущность объекта, выйти за пределы индивидуального мира. На его основе формируется человеческое знание. Оно передается из поколения в поколение посредством записи словесных или знаковых сигналов.
Правое полушарие осуществляет образное мышление. Оперируя образами объектов внешнего мира, оно может создавать из них небывалые, фантастические комбинации. А это - основа творчества, принятия необычных решений. Исключительно велико значение правого полушария для музыкального и художественного творчества. Известно, что наиболее выдающиеся художники, поэты, музыканты - это люди с преобладанием правополушарного мышления.
Несмотря на функциональную асимметрию, мозг работает как единое целое. Обобщая информацию, он обеспечивает адекватное поведение, мышление, сознание, память, трудовую и творческую деятельность человека.



