Funkce přední části mozku. Mozek
Mozek, mozek, je umístěn v lebeční dutině a má tvar v obecný obrys odpovídající vnitřním obrysům lebeční dutiny. Jeho superolaterální neboli dorzální povrch je v souladu s lebeční klenbou konvexní a ventrální strana neboli základna mozku je víceméně zploštělá a nerovná. Mozek lze rozdělit na tři velké části: hemisféry velký mozek(hemispheriae cerebri), mozeček (cerebellum) a mozkový kmen (truncus cerebri). Největší část celého mozku zabírají hemisféry, co do velikosti následuje mozeček, zbývající, relativně malou část tvoří mozkový kmen.
mozek. Obě hemisféry jsou od sebe odděleny trhlinou, fisstira longitudinalis cerebri, jdoucí sagitálním směrem. V hloubce podélné trhliny jsou hemisféry navzájem spojeny adhezemi - corpus callosum, corpus callosum a další podkladové formace. Před corpus callosum je podélná štěrbina průchozí a za ní přechází do příčné štěrbiny mozku, fissiira transversa cerebri, oddělující zadní části hemisfér od základního mozečku.
Spodní povrch mozku(obr. 272). Ze strany spodní plochy mozku bledne nižší mozek, je viditelná nejen spodní strana mozkových hemisfér a mozečku, ale i celá spodní plocha mozkového kmene a také nervy vybíhající z mozku. Zde se setkáme s následujícími částmi, půjdeme-li zepředu dozadu. Přední část spodního povrchu mozku představují čelní laloky hemisfér. Na spodní ploše jsou patrné čelní laloky čichové žárovky, btilbi olfactorii, ke kterým z nosní dutiny přistupují tenká nervová vlákna, fila olfactoria, otvory lamina cribrosa ethmoidální kosti, které společně tvoří první pár hlavových nervů - čichové nervy, nn. olfactorii. Obvykle se při vyjímání mozku z lebky tyto závity odtrhávají od billbus olfactorius. Čichové bulby pokračují zezadu dovnitř čichové dráhy, tractus olfactorii, z nichž každý končí dvěma kořeny, mezi nimiž je vyvýšení tzv trigonum olfactorium. Přímo za tím druhým na obou stranách je přední perforovaná látka, substantia perforata anterior, tak pojmenovaný kvůli malým otvorům, které se zde nacházejí, kterými procházejí krevní cévy do dřeně.
Uprostřed mezi oběma předními perforovanými prostory leží optický chiasma, chiasma opticum ve tvaru písmene "X". Z horního povrchu chiasmatu vyčnívá tenká deska šedá, lamina terminalis, jdoucí hlouběji fissura longitudinis cerebri. Umístěno za optickým chiasmatem šedý hrbolek, tuber cinereum; její vrchol je protáhlý v úzkou trubku, t. zv trychtýř, infundibulum, na kterém, jako bobule na stopce, visí kulaté tělo - přívěsek mozku, hypophysis cerebri. Když je mozek umístěn na své místo v lebce, přívěsek je umístěn do vybrání sella turcica. Za šedou mohylou jsou dvě kulovité bílý nadmořské výšky - mastoidní těla, corpora mamillaria. Za nimi leží poměrně hluboká čtyřúhelníková jáma, fossa interpeduncularis, ohraničený bočně dvěma tlustými bílými chomáčemi sbíhajícími se dozadu a tzv nohy mozku, pedunculi cerebri. Dno jámy je provrtáno otvory pro nádoby, a proto je tzv zadní perforovaná látka, substantia perforata posterior. Vedle této látky, v drážce mediálního okraje mozkové stopky, vystupuje na obou stranách třetí pár - okulomotorický nerv, n. oculomotorius. Nejtenčí z mozkových nervů je viditelný na straně mozkových stopek trochleární nerv, n. trochlearis- IV pár, který však nevzniká ze spodiny mozku, ale z jeho dorzální strany, z tzv. horního medulárního velum. Za mozkovými stopkami je tlustý příčný dřík - most, pons(Varolii), který se zužuje do strany a noří se do mozečku. Boční části mostu nejblíže k mozečku se nazývají střední cerebelární stopka, pedunculi cerebellares medii; na hranici mezi nimi a samotným mostem se na obou stranách objevuje pár V - trojklaného nervu, n. trigeminus. Za mostem leží medulla, prodloužená medulla; mezi ním a zadním okrajem můstku po stranách střední čáry je viditelný začátek VI páru - abdukuje nerv, n. abducens; ještě více stranou, u zadního okraje středních mozečkových stopek, vystupují vedle sebe na obou stranách další dva nervy: VII pár - obličejový nerv, n. facialis, a VIII pár - n. vestibulocochlearis.
Mezi pyramidou a olivovou medulla oblongata vystupují kořeny páru XII - hypoglossální nerv, n. hypoglossus. Páry kořenů IX, X a XI - n. glossopharyngeus, n. vagus a n. příslušenství(horní část) - vystupují z drážky za olivou. Spodní vlákna páru XI již odcházejí z míchy v její krční části.
Vývoj (embryogeneze) mozku
Dřeňová trubice je velmi brzy rozdělena na dvě části odpovídající mozku a míše. Jeho přední, rozšířená část, představující rudiment mozku, jak již bylo zmíněno, je rozřezána zúžením na tři primární medulární váčky ležící za sebou: přední, prosencephalon, střední, mezimozek a zadní, rombencefalon. Přední mozkový váček je vpředu uzavřen tzv. koncovou ploténkou, lamina terminalis. Toto stadium tří váčků se po následné diferenciaci změní na pět váčků, z nichž vznikne pět hlavních částí mozku (obr. 273). Současně se mozková trubice ohýbá v sagitálním směru. Nejprve se v oblasti středního váčku rozvine parietální křivka konvexní k dorzální straně a poté na hranici s rudimentem míchy také okcipitální křivka konvexně dorzálně. Mezi nimi se v oblasti zadního váčku vytvoří třetí ohyb, konvexní k ventrální straně (pontinní ohyb).
Přes tuto poslední zatáčku zadní medulární váček, rhombencephalon, je rozdělena do dvou částí. Z nich se zadní část, myelencephalon, během konečného vývoje přeměňuje v prodlouženou míchu a z přední části, zvané fencephalon, se na ventrální straně vyvíjí mostík a na dorzální straně cerebellum. Metencephalon je oddělen od vezikuly středního mozku ležící před ním úzkým zúžením, isthmus rhombencephali. Společná dutina rhombencephalon, která má ve frontální části tvar kosočtverce, tvoří IV komoru, komunikující s centrálním kanálem míšním. Jeho ventrální a laterální stěny v důsledku vývoje jader hlavových nervů v nich značně zesílí, zatímco dorzální stěna zůstává tenká. V oblasti prodloužené míchy je většina tvořena pouze jednou epiteliální vrstvou, srostlou s cévnatkou (tela chorioidea inferior). střední bublina, mezimozek, houstnou s vývojem mozkové hmoty v nich rovnoměrněji. Ventrálně z nich vycházejí mozkové stopky a na dorzální straně - quadrigeminální deska (viz obr. 273). Dutina střední vezikuly se mění v úzký kanál - akvadukt, který se připojuje k IV komoře.
prochází výraznější diferenciací a úpravami ve formě přední medulární váček, prosencephalon, která se dělí na zadní část, diencephalon (diencephalon) a přední část, telencephalon ( telencephalon). Boční stěny diencefala, ztluštění, tvoří zrakové tuberkuly (thalami). Boční stěny, vyčnívající do stran, navíc tvoří dva optické váčky, ze kterých se následně vyvíjí sítnice a zrakové nervy. Hřbetní stěna diencephalonu zůstává tenká, ve formě epiteliální destičky srostlé s cévnatkou (tela chorioidea superior). Vzadu z této stěny vzniká výběžek, díky kterému dochází k epifýze (corpus pineale). Duté nohy optických váčků jsou zataženy z ventrální strany do stěny přední mozkové váčky, v důsledku čehož se na dně dutiny posledně jmenovaného vytvoří prohlubeň, recessus opticus, z níž se skládá přední stěna tenké lamina terminalis. Za recessus opticus se objevuje další trychtýřovitá prohlubeň, z jejíchž stěn vzniká tuber cinereum, infundibulum a zadní (nervový) lalok hypophysis cerebri. Ještě dále vzadu, v oblasti diencephalonu, se tvoří párové corpora mamillaria ve formě jediné elevace. Dutina diencephalonu tvoří třetí komoru.
Telencephalon se dělí na střední, menší část (telencephalon medium) a dvě velké postranní části – váčky mozkových hemisfér (hemispherium dextrum et sinistrum), které u člověka rostou velmi silně a na konci vývoje jejich velikost výrazně převyšuje zbytek mozku. Dutina telencephalon media, která je předním pokračováním dutiny diencephalon (komora III), komunikuje po stranách přes mezikomorové otvory s dutinami váčků hemisfér, které jsou na vyvinutý mozek jsou nazývány postranní komory. Přední stěna střední části telencephalon media, která je přímým pokračováním lamina terminalis, vytváří na začátku prvního měsíce embryonálního života ztluštění, tzv. komisurální ploténku, ze které corpus callosum a přední komisura.
Na bázi váčků hemisfér je uvnitř na obou stranách vytvořen výběžek, t. zv. nodulární tuberkul, ze kterého se vyvíjí striatum, corpus striatum. Část mediální stěny vezikuly hemisfér zůstává ve formě jediné epiteliální vrstvy, která je do váčku svinuta záhybem cévnatky (plexus chorioideus). Na spodní straně každého vezikula hemisféry se již v 5. týdnu embryonálního života vyskytuje výběžek - rudiment čichový mozek, rhinencephalon, která je postupně od stěny hemisfér vymezována rýhou odpovídající fissura rhinalis lateralis. S rozvojem šedé hmoty (kortexu) a následně bílé hmoty ve stěnách hemisféry se tato zvětšuje a tvoří tzv. plášť, pallium, ležící nad čichovým mozkem a pokrývající nejen zrakové pahorky, ale i dorzální povrch středního mozku a mozečku.
Jak roste hemisféra, zvětšuje se nejprve ve frontálním laloku, poté v parietálním a týlním laloku a nakonec ve spánkovém laloku. Díky tomu to vypadá, jako by se plášť otáčel kolem zrakových cípů, nejprve zepředu dozadu, pak dolů a nakonec se zakřivil dopředu směrem k čelnímu laloku. Výsledkem je, že na boční ploše polokoule, mezi čelním lalokem a spánkovým lalokem, který se k němu přibližuje, se vytvoří otvor, fossa cerebri lateralis (Sylvii), který, když se jmenované mozkové laloky úplně spojí, otočí do mezery, sulcus cerebri lateralis (Sylvii); na jeho dně vzniká ostrůvek, insula.
Během vývoje a růstu hemisféry spolu s ní naznačená „rotace“ a její vnitřní komory, laterální komory mozku (zbytky dutiny primárního močového měchýře), jakož i část corpus striatum (caudatum). nucleus) vyvinout a provést naznačenou „rotaci“, která vysvětluje podobnost jejich tvaru s tvarem hemisféry: komory mají přední, střední a zadní část a spodní část, která se zakřivuje dolů a dopředu (viz obr. 295) caudate nucleus má hlavu, tělo a ocas, které se zakřivují dolů a dopředu.
Brázdy a konvoluce(obr. 274, 275, 276) vznikají v důsledku nerovnoměrného růstu samotného mozku (D. Zernov), který souvisí s vývojem jeho jednotlivých částí. Tak se na místě čichového mozku objevují sulcus olfactorius, sulcus hyppocdmpi a sulcus cinguli; na hranici kortikálních konců kožního a motorického analyzátoru (koncepce analyzátoru a popis rýh viz níže) - sulcus centralis; na rozhraní motorického analyzátoru a premotorické zóny, která přijímá impulsy z viscera - sulcus precentralis; místo sluchového analyzátoru - sulcus temporalis superior; v oblasti vizuálního analyzátoru - sulcus calcarinus a sulcus parieiooccipitdlis.
Všechny tyto brázdy, které se objevují dříve než jiné a vyznačují se naprostou stálostí, patří podle D. Zernova k brázdy první kategorie. Zbývající drážky, které mají jména a také vznikají v souvislosti s vývojem analyzátorů, ale objevují se o něco později a jsou méně konstantní, patří mezi brázdy druhé kategorie. V době narození jsou přítomny všechny brázdy první a druhé kategorie. A konečně, četné malé rýhy, které nemají jména, se objevují nejen v životě dělohy, ale také po narození. Jsou extrémně proměnlivé v době výskytu, místě a počtu; Tento brázdy třetí kategorie. Rozmanitost a složitost mozkového reliéfu závisí na stupni jejich rozvoje. Výška lidský mozek v embryonálním období a v prvních letech života, kdy tělo rychle roste, adaptuje se na nové prostředí, osvojuje si schopnost vzpřímené chůze a vytváří druhou, verbální, signalizační systém, probíhá velmi intenzivně a končí ve věku 20 let. U novorozenců váží mozek (v průměru) 340 g u chlapců a 330 g u dívek a u dospělého 1375 g u mužů a 1245 g u žen.
Části mozku
Na základě embryonálního vývoje, jak již bylo naznačeno, je mozek rozdělen na části umístěné od kaudálního konce v následujícím pořadí:
1) rhombencephalon, kosočtverec nebo zadní mozek, který se zase skládá z: a) myelencephalon, prodloužené míchy a b) metencephalon - samotný zadní mozek; 2) mezimozek- střední mozek; 3) prosencephalon, přední mozek, ve kterém rozlišují: a) diencephalon - diencephalon a b) telencephalon - telencephalon.
Kromě těchto úseků se mezi kosočtvercem a středním mozkem nachází ještě isthmus, isthmus rhombencephali.
Mezimozek a prosencefalon tvoří velký mozek, velký mozek, na rozdíl od malého mozku mozeček – mozeček.
Rhombencephalon
Myelencephalon, medulla oblongata
Medulla oblongata, tnyelencephalon, medulla oblongdta(obr. 277, 278), představuje přímé pokračování míchy do mozkového kmene a je součástí kosočtverce neboli zadního mozku. Kombinuje strukturální rysy míchy a počáteční části mozku, což ospravedlňuje jeho název - myelencephalon. Medulla oblongata má vzhled cibule, bulbus cerebri, s. bulbus medullae spinalis (odtud termín „bulvární poruchy“); horní rozšířený konec ohraničuje most a jeho spodní hranice je výstupním bodem kořenů prvního páru krčních nervů nebo úrovní foramen magnum týlní kosti.


1. Na přední (ventrální) ploše Fissura mediana anterior probíhá podél střední linie prodloužené míchy a tvoří pokračování stejnojmenné míšní rýhy. Na jeho stranách, na obou stranách, jsou dva podélné prameny - pyramidy, pyramides medullae oblongatae, které jakoby pokračují do předních provazců míšních. Svazky nervových vláken tvořících pyramidu se částečně protínají v hloubce fissura mediana anterior s podobnými vlákny opačné strany - decussidtio pyramidum, načež sestupují v postranním provazci na druhé straně míchy - tractus corticospinal (pyramidalis) lateralis, částečně zůstávají nezkřížené a sestupují v předním provazci míchy na její straně - tractus kortikospinální (pyramidalis) anterior.
Pyramidy chybí u nižších obratlovců a objevují se, když se neokortex vyvíjí; proto jsou nejrozvinutější u lidí, neboť pyramidální vlákna spojují mozkovou kůru, která dosáhla svého nejvyššího rozvoje u lidí, s jádry mozkových nervů a předními rohy míšními.
Bočně k pyramidě leží oválná vyvýšenina – olivová, olfva, která je od pyramidy oddělena žlábkem, sulcus lateralis anterior.
2. Na hřbetní (zadní) straně Medulla oblongata (viz obr. 278) se táhne sulcus medidnus posterior - přímé pokračování stejnojmenné míšní rýhy. Po jeho stranách leží zadní provazce, ohraničené laterálně na obou stranách slabě definovaným sulcus lateralis posterior. Směrem nahoru se zadní provazce rozbíhají do stran a jdou do mozečku a stávají se součástí jeho dolních končetin, pedunculi cerebellares inferiores, ohraničující kosočtverec dole. Každý zadní provazec je rozdělen střední rýhou na střední, fasciculus gracilis a boční, fasciculus cuneatus. V dolním rohu kosočtverečné fossa získávají jemné a klínovité provazce ztluštění - tuberculum nuclei gracilis a tuberculum nuclei cunedti. Tato ztluštění jsou způsobena jádry šedé hmoty, nucleus gracilis a nucleus cuneatus, které jsou podobné provazcům. V těchto jádrech končí vzestupná vlákna dorzálních kořenů míchy (svazky Gaulle a Burdach) procházející v zadních provazcích. Boční povrch medulla oblongata, který se nachází mezi sulcus lateralis posterior et anterior, odpovídá postrannímu provazci. XI, X a IX páry hlavových nervů vycházejí ze sulcus lateralis posterior za olivou. Medulla oblongata obsahuje Spodní část kosočtverec fossa (obr. 279; viz obr. 283).

Vnitřní struktura medulla oblongata. Dřeň prodloužená vznikla v souvislosti s rozvojem statických a akustických orgánů a také v souvislosti s žaberním aparátem souvisejícím s dýcháním a krevním oběhem. Proto obsahuje jádra šedé hmoty, související s rovnováhou a koordinací pohybů a také s regulací metabolismu (obr. 280).
1. Nucleus olivdris, olivové jádro, má vzhled svinuté desky šedé hmoty, otevřené mediálně (hilus) a způsobuje vyčnívání olivy zvenčí. Je spojena s dentátním jádrem mozečku a je středním balančním jádrem, nejvýraznějším u člověka, jehož vertikální poloha vyžaduje nejvyspělejší rovnovážný aparát. (Existuje také přístup nucl. olivaris, med.)
2. Formatio reticularis, retikulární útvar, vytvořený propletením nervových vláken a nervových buněk ležících mezi nimi.
3. Jádra čtyř párů dolních hlavových nervů (XII-IX), související s inervací derivátů branchiálního aparátu a vnitřností.
4. Vitální centra dýchání a oběhu spojená s jádry bloudivého nervu. Pokud tedy dojde k poškození prodloužené míchy, může nastat smrt.
bílá hmota Medulla oblongata obsahuje dlouhá a krátká vlákna. Dlouhé zahrnují sestupné pyramidové dráhy, které procházejí předními provazci míchy a částečně se protínají v oblasti pyramid. Navíc z jader zadních funiculi (nucleus gracilis et cuneatus) začínají druhé neurony vzestupných smyslových drah, jdoucí z medulla oblongata do zrakového thalamu, trdctus bulboihaldmicus. Vlákna tohoto svazku tvoří střední smyčku, lemniscus medlis, která se kříží v prodloužené míše, decussedtio lemniscorum, a ve formě svazku vláken umístěného dorzálně k pyramidám mezi olivami - vrstva interolivové smyčky - jde dále. V prodloužené míše jsou tedy dvě křížení dlouhých drah: ventrální motorická, decussatio pyramidum, a dorzální senzorická, decussatio lemniscorum.
Krátké dráhy zahrnují svazky nervových vláken, které spojují jednotlivá jádra šedé hmoty a také jádra prodloužené míchy se sousedními částmi mozku. Mezi nimi stojí za zmínku trdctus olivocerebellars a fasciculus longitudindlis medlis ležící dorzálně k interolivové vrstvě.
Topografické vztahy hlavních útvarů medulla oblongata jsou patrné na řezu pořízeném na úrovni oliv (viz obr. 280). Kořeny vycházející z jader hypoglossálního a vagusového nervu rozdělují prodlouženou míchu na obou stranách do tří oblastí: zadní, boční a přední. V zadní části jsou jádra zadního funiculu a dolních cerebelárních stopek, v laterálních jsou olivové jádro a formatio reticularis a v přední části jsou pyramidy.
Metencephalon
Metencephalon se skládá ze dvou částí: ventrální - most pons a dorzální - mozeček.
Most
Most, most(Varolii), je tlustá bílá šachta na spodině mozku, hraničící kaudálně s horním koncem prodloužené míchy a kraniálně s mozkovými stopkami (viz obr. 277). Laterální hranice můstku je uměle nakreslená čára přes kořeny trigeminálního a lícního nervu, linea trigeminofacidlis. Po straně této linie jsou střední cerebelární stopky, pedunculi cerebellares medii, nořící se na obě strany do mozečku. Hřbetní plocha ponsu není zvenčí viditelná, protože je skryta pod mozečkem, který tvoří horní část kosočtverečné jamky (podlaha čtvrté komory). Ventrální povrch můstku má vláknitou povahu a vlákna obecně probíhají příčně a směřují do pedunculi cerebellares medii. Podél střední linie ventrální plochy je mírná rýha, sulcus basilaris, ve které leží a. basilaris.
Vnitřní konstrukce mostu. Na příčných řezech mostu je vidět, že se skládá z většího spodního, popř ventrální část, pars ventralis pontis a menší hřbetní, pars dorsalis pontis. Hranici mezi nimi tvoří silná vrstva příčných vláken – lichoběžníkové tělísko, corpus trapezoideum, jehož vlákna patří do sluchové dráhy. V oblasti lichoběžníkového těla se nachází jádro, které souvisí i se sluchovou dráhou – nucleus dorsalis corporis trapezoidei, neboli nucleus olivaris superior (BNA). Posledně jmenované jméno bylo dáno jádru, protože má zubatý tvar, podobný jádru prodloužené míchy olivary.
Pars ventralis obsahuje podélná a příčná vlákna, mezi kterými jsou rozptýlena vlastní jádra šedé hmoty, nuclei pontis.
Podélná vlákna patří do pyramidálních drah, fibrae corticopontinae, které jsou napojeny na vlastní jádra mostu, odkud vycházejí příčná vlákna směřující do kůry mozečku, tractus pontocerebellaris. Celý tento systém drah spojuje mozkovou kůru s kůrou mozečku přes most. Čím vyvinutější je mozková kůra, tím vyvinutější je most a mozeček. Most se přirozeně ukazuje být nejvýraznější u lidí, což je specifická vlastnost struktura jeho mozku. V pars dorsalis je formatio reticularis pontis, který je pokračováním stejné části prodloužené míchy, a na vrcholu retikulární formace je dno kosočtverečné jamky lemované ependymem s podložními jádry hlavových nervů ( VIII-V páry).
V pars dorsalis pokračují také vodivé dráhy prodloužené míchy, umístěné mezi střední linií a nucleus dorsalis corporis trapezoidei a částí mediální kličky, lemniscus medialis; v posledně jmenovaném se protínají vzestupné dráhy medulla oblongata, tractus bulbothalamicus.
Mozeček
Mozeček, mozeček, je derivátem zadního mozku, který se vyvinul v souvislosti se statickými receptory. Proto přímo souvisí s koordinací pohybů a je orgánem adaptace těla na překonání základních vlastností tělesné hmoty - gravitace a setrvačnosti. Je také považován za jedno z nejvyšších center autonomního (sympatického) nervového systému (L. A. Orbeli a jeho škola).
Vývoj mozečku v procesu fylogeneze prošel 3 hlavními fázemi, odpovídajícími změnám ve způsobech pohybu zvířete.
V první fázi, kdy se živočichové ve vodě pohybovali hadovitými pohyby těla (cyklostomy), vypadal jejich mozeček jako malý útvar, auricula, který se u suchozemských živočichů vyvinul v tzv. vločky. Jedná se o nejstarší část mozečku - archicerebellum. Ve druhé fázi, kdy se u vodních živočichů objevují ploutve (ryby) a mění se pohyby trupu, se objevuje nová část mozečku - vermis, což je u lidí stará část mozečku, paleocerebellum. Konečně ve třetí fázi se u suchozemských zvířat objevují mozečkové hemisféry, což je nejnovější část mozečku, neocerebellum, a postupně rostou, jak se zvířecí tělo vzdaluje od země a vyvíjí se lokomoce s končetinami. U lidí díky vzpřímené chůzi za pomoci jednoho páru končetin (nohy) a zlepšení uchopovacích pohybů ruky s pracovní procesy dosahují mozečkové hemisféry největší rozvoj, takže mozeček u lidí je vyvinutější než u všech zvířat, což je specifikum lidská vlastnost jeho budovy.
Mozeček se nachází pod okcipitálními laloky mozkových hemisfér, dorzálně k mostu a prodloužené míše a leží v zadní lebeční jámě. Má objemné boční díly, popř hemisféry, hemispheria cerebelli, a střední úzká část umístěná mezi nimi - červ, vermis.
Na předním okraji cerebellum je přední zářez, který obklopuje přilehlou část mozkového kmene. Na zadním okraji je užší zadní zářez, který odděluje hemisféry od sebe.
Povrch mozečku je pokryt vrstvou šedé hmoty, která tvoří kůru mozečku a tvoří úzké konvoluce, folia cerebelli, oddělené od sebe rýhami, fissurae cerebelli. Mezi nimi nejhlubší, fissura horizontdlis cerebelli, probíhá podél zadního okraje mozečku a odděluje horní povrch hemisfér, fdcies superior, od spodní, fdcies inferior. Pomocí vodorovných a jiných velkých rýh je celý povrch mozečku rozdělen na řadu lalůčků, lobuli cerebelli. Mezi nimi je třeba vyzdvihnout nejvíce izolovaný malý lalok - vločku, ležící na spodní ploše každé hemisféry u středního mozečkového stopky, stejně jako část vermis spojenou s vločkou - nodulus, nodulus. Floculus je spojen s nodulem pomocí tenkého proužku - stopka flocculus, pedunculus flocculi, která mediálně přechází v tenkou poloměsíčitou ploténku - velum dřeňové inferior, velum medullare inferius.
Vnitřní struktura mozečku. V tloušťce mozečku jsou párová jádra šedé hmoty, umístěná v každé polovině mozečku, mezi jeho bílou hmotou (obr. 281). Po stranách střední čáry, v oblasti, kde stan, fastigium, vyčnívá do mozečku, leží nejstřednější jádro - jádro stanu, nucleus fasiigii. Bočně k němu jsou malé ostrůvky sférického jádra, nucleus globosus, a ještě více laterálně - korkové jádro, nucleus emboliformis. Konečně ve středu hemisféry je zubaté jádro, nucleus denttus, které vypadá jako šedá vinutá destička, podobná olivovému jádru, a mediálně otevřené - hilus nuclei dentati. Podobnost mezi nucleus dentatus mozečku a olivárním jádrem, které je také zubaté, není náhodná, protože obě jádra jsou spojena drahami, fibrae olivocerebellares, a každý gyrus jednoho jádra je podobný gyru druhého. Obě jádra se tedy společně podílejí na realizaci rovnovážné funkce (viz obr. 280, 281).
Jmenovaná cerebelární jádra mají různá fylogenetická stáří: nucleus fastigii označuje nejstarší část mozečku - flocculus (archicerebellum), spojený s vestibulárním aparátem, nuclei emboliformis et globosus - starou část (paleocerebellum), která vznikla v souvislosti s pohyby těla, a nucleus dentatus - k nejmladšímu (neocerebellum), který se vyvinul v souvislosti s pohybem pomocí končetin. Proto, když je každá z těchto částí poškozena, různé strany motorická funkce, odpovídající různým stádiím fylogeneze, a to: při poškození flokulonodulárního systému a jeho stanového jádra dochází k narušení rovnováhy těla. Při poškození červa a jeho odpovídajících korkových a kulovitých jader je narušeno fungování svalů krku a trupu. Při postižení hemisfér a zubatého jádra pracují svaly končetin.
Bílá hmota mozečku na řezu má vzhled malých listů rostliny, odpovídajících každému gyrusu, pokrytých na periferii kůrou šedé hmoty. Výsledkem je, že celkový obraz bílé a šedé hmoty na části mozečku připomíná strom, arbor viiae cerebelli (strom života; pojmenovaný po vzhled, protože poškození mozečku nepředstavuje bezprostřední ohrožení života). Bílá hmota mozečku je složena z různých typů nervových vláken. Některé z nich spojují gyri a lalůčky, jiné jdou z kůry do vnitřních jader mozečku a nakonec další spojují mozeček se sousedními částmi mozku. Tato poslední vlákna jsou zahrnuta v tři páry cerebelárních stopek:
1. Bérce, pedunculi cerebellares inferiores(k prodloužené míše).
Patří mezi ně trdctus spinocerebelldris posterior (Flechsig) směřující do mozečku, fibrae arcudtae externae - z jader zadních provazců medulla oblongata a fibrae olivocerebellars - z olivy. Všechna tato vlákna končí v kůře červa a hemisférách. Kromě toho existují vlákna z jader vestibulárního nervu, končící v nucleus fastigii. Díky všem těmto vláknům přijímá mozeček impulsy z vestibulárního aparátu a proprioceptivního pole, v důsledku čehož se stává jádrem proprioceptivní senzitivity a provádí automatické korekce motorické aktivity zbytku mozku. Součástí bérců jsou i sestupné dráhy v opačném směru, a to: z nucleus fastigii do laterálního vestibulárního jádra a z něj do předních rohů míšních, tractus vestibulospinalis. Prostřednictvím této cesty mozeček ovlivňuje míchu.
2. Střední nohy, pedunculi cerebellares medii(na most).
Obsahují nervová vlákna od pontinních jader až po kůru mozečku. Dráhy do mozečkové kůry, trdctus pontocerebellars, které vznikají v pontinních jádrech, jsou umístěny v pokračování kortikálno-pontinních drah, fibrae corticopontinae, po dekusaci končí v pontinních jádrech. Tyto dráhy spojují mozkovou kůru s kůrou mozečku, což vysvětluje skutečnost, že čím vyvinutější mozková kůra, tím vyvinutější je mostík a mozkové hemisféry, což je pozorováno u lidí.
3. Horní končetiny, pedunculi cerebellares superiores(do quadrigeminu). Skládají se z nervových vláken probíhajících v obou směrech: 1) do mozečku - trdctus spinocerebelldris anterior (Gowers) a 2) od nucleus dentatus cerebellum ke stropu quadrigeminalis - trdctus cerebellotegmentdlis, který po dekusaci končí červeném jádře a v optickém thalamu. Prvními drahami přijímá mozeček impulsy z míchy a druhou vysílá impulsy do extrapyramidového systému, kterým sám míchu ovlivňuje.
Šíje
Isthmus, isthmus rhombencephali, představuje přechod z rhombencefala do mezencefala. Skládá se z: 1) horních cerebelárních stopek, pedunculi cerebellares superiores; 2) horní medulární velum natažené mezi nimi a cerebellum, velum medullare superius, který je připojen ke střední rýze mezi kvadrigeminálními tuberkulami; 3) smyčkový trojúhelník, trigonum lemnisci, způsobené průběhem sluchových vláken laterálního lemniscus, lemniscus lateralis. Tento šedý trojúhelník je ohraničen vpředu spodní rukojetí quadrigeminu, vzadu horním mozečkovým stopkou a laterálně stopkou mozkovou. Ten je oddělen od isthmu a středního mozku jasně definovanou rýhou, sulcus lateralis mesencephali. Horní konec čtvrté komory vyčnívá do isthmu a přechází do akvaduktu ve středním mozku.
IV komora
IV komora, ventricuius quartus, je pozůstatkem dutiny zadního mozku a je tedy společnou dutinou pro všechny části zadního mozku, které tvoří kosočtverec (medulla oblongata, cerebellum, pons a isthmus). IV komora připomíná stan, ve kterém se rozlišuje dno a střecha (viz obr. 279; obr. 282).
Dno, nebo základna, komory má tvar kosočtverce, jako by byl vtlačen do zadní plochy prodloužené míchy a mostu. Proto mu říkají kosočtverec fossa, fossa rhomboidea. Otevírá se do posteroinferiorního rohu kosočtverečné jamky středový kanál míchu a v anterosuperiorním rohu IV komory komunikuje s akvaduktem. Boční úhly končí slepě ve formě dvou kapes, recessus laterdles ventriculi qudrti, zakřivených ventrálně kolem dolních mozečkových stopek.
Střecha IV komory, tegmen ventriculi quarti, má tvar stanu, fastigium (BNA), a skládá se ze dvou mozkových plachet: horní, velum medullare superius, natažená mezi horními cerebelárními stopkami, a spodní, velum medullare inferius, párový útvar sousedící s stopky mozečku. Část střechy mezi plachtami je tvořena hmotou mozečku. Spodní medulární velum je doplněno vrstvou cévnatky tela chorioidea ventriculi qudrti, pokrytou zevnitř vrstvou epitelu, lamina chorioidea epithelialis, představující rudiment zadní stěny zadního medulárního měchýře (plexus je spojen s it - pl. cévnatka. ventr. IV).
Tělo chorioidea zpočátku úplně uzavírá dutinu komory, ale pak se v ní během vývoje objeví tři otvory: jeden v oblasti spodního úhlu kosočtverečné jamky, apertura medina ventriculi qudrti (foramen Magendi) (největší), a dvě v oblasti laterálních vybrání komory, aperturae laterdles ventriculi qudrti (foramina Luschka). Těmito otvory čtvrtá komora komunikuje se subarachnoidálním prostorem mozku, díky čemuž mozkomíšní mok proudí z mozkových komor do intertekálních prostor. V případě zúžení nebo srůstu těchto otvorů v důsledku zánětu mozkových blan (meningitidy) nenachází mozkomíšní mok hromadící se v mozkových komorách vývod do subarachnoidálního prostoru a dochází k mozkové kapavce.
Diamantová fossa, fossa rhomboidea(obr. 283), má čtyři strany podle tvaru kosočtverce - dvě horní a dvě spodní. Horní strany kosočtverce jsou ohraničeny dvěma horními cerebelárními stopkami a spodní strany dvěma spodními stopkami. Podél kosočtverce, podél střední čáry, od horního k dolnímu rohu je střední rýha, sulcus medidnus, která rozděluje kosočtvercovou jamku na pravou a levou polovinu. Po stranách brázdy je dvojice vyvýšení, eminentia medialis způsobené nahromaděním šedé hmoty.
V dolní části se eminentia medialis postupně zužuje, přechází v trojúhelník, na který se promítá jádro n. hypoglossalis, trigonum nervi hypoglosi. Bočně ke spodní části tohoto trojúhelníku leží menší trojúhelník, který je patrný svou šedou barvou, trigonum n. vagi, který obsahuje autonomní jádro nervu vagus, nucleus dorsdlis nervi vagi. Nahoře má eminentia medialis elevaci - tuber obličeje, collicuius facialis, způsobený průchodem kořene obličeje a výběžkem n. nucleus abducens.
V oblasti bočních úhlů se na obou stranách nachází vestibulární pole, area vestibularis. Zde jsou umístěna jádra páru VIII. Některá vlákna z nich vycházející probíhají napříč kosočtvercovou jamkou z laterálních úhlů ke střednímu sulcus ve formě vodorovných pruhů, striae medulldres ventriculi qudrti. Tyto pruhy rozdělují rhomboid fossa na horní a dolní polovinu a odpovídají hranici mezi prodlouženou míchou a mostem.
Topografie šedé hmoty kosočtverečné jamky. Šedá hmota míchy přímo přechází do šedé hmoty mozkového kmene a částečně se šíří podél kosočtverečné jamky a stěn akvaduktu (viz střední mozek) a částečně je rozdělena na samostatná jádra mozkových nervů nebo jádra svazků cest.
Abychom pochopili umístění těchto jader, je třeba vzít v úvahu, jak bylo uvedeno výše, že uzavřená dřeňová trubice míchy se při přechodu do prodloužené míchy na své zadní straně otevřela a rozvinula se do kosočtverečné jamky. V důsledku toho se zdálo, že se zadní rohy šedé hmoty míšní rozcházejí do stran. Somaticko-senzitivní jádra umístěná v zadních rozích byla umístěna laterálně v kosočtverečné jámě a somatomotorická jádra odpovídající předním rohům zůstala umístěna mediálně. Pokud jde o vegetativní jádra umístěná v postranních rozích míšních, pak se podle polohy postranních rohů mezi zadním a předním ukázalo, že tato jádra, když se mozková trubice rozšířila, ležela v kosočtverečné jamce mezi somaticko-senzitivní a somaticko-motorická jádra. V důsledku toho se ve stěně kosočtverečné jamky na rozdíl od míchy jádra šedé hmoty nenacházejí v předozadním směru, ale leží v řadách - mediálně a laterálně.
Takže například somaticko-motorická jádra párů XII a VI leží v mediální řadě, vegetativní jádra párů X, IX, VII jsou ve střední řadě a somaticko-citlivá jádra páru VIII jsou postranní.
Projekce jader mozkových nervů do kosočtverečné jamky:
XII pár - hypoglosální nerv, n. hypoglossus, má jediné motorické jádro umístěné v nejnižší části kosočtverečné jamky, v hloubce trigona n. hypoglosy.
XI pár - přídatný nerv, n. příslušenství, má dvě jádra (obě motorické): jedno se nachází v míše a nazývá se nucleus spinalis n. accessorii, druhý je kaudálním pokračováním jader párů X a IX nervů a nazývá se nucleus ambiguus. Leží v medulla oblongata dorzolaterálně k jádru olivy.
X pár - bloudivý nerv, n. vagus, má tři jádra:
1. Citlivé jádro, nucleus trdctus solitdrii, se nachází vedle jádra n. hypoglossalis hluboko v trigonu n. Vagi.
2. Vegetativní jádro, nucleus dorsalis i. vagi, leží ve stejné oblasti.
3. Motorické jádro, nucleus ambiguus (double) je společné s jádrem páru IX, umístěným ve formatio reticularis, hlouběji než nucleus dorsalis.
IX pár - n. glosofaryngeální, n. glossopharyngeus, také obsahuje tři jádra:
1. Citlivé jádro, nucleus trdctus solitdrii, leží laterálně od jádra n. hypoglossus.
2. Vegetativní (sekreční) jádro, nucleus salivatorius inferior, dolní slinné jádro; její buňky jsou roztroušeny ve formacio reticularis medulla oblongata mezi n. ambiguus a olivové jádro.
3. Motorické jádro, společné s n. vagus a n. accessorius, nucleus ambiguus.
VIII pár - staticko-sluchový nerv, n. vestibulocochlearis, má mnohočetná jádra vyčnívající do laterálních rohů kosočtverečné fossa, v oblasti vestibularis. Jádra jsou rozdělena do dvou skupin, odpovídajících dvěma částem nervu; jedna část nervu, pars cochlearis - kochleární nerv, nebo samotný sluchový nerv, má dvě jádra: dorzální, nucleus cochleans dorsdlis, a ventrální, nucleus cochlearis ventrdlis, umístěné laterálně a před předchozím. Další část nervu, pars vestibularis - nerv vestibulu, neboli statický nerv, má čtyři jádra (nuclei vestibulares):
1. Mediální - hlavní (Schwalbeho jádro).
2. Laterální - Deitersovo jádro.
3. Horní - Bechtěrevovo jádro.
4. Spodní.
Přítomnost čtyř jader u lidí odráží raná stádia fylogeneze, kdy ryby měly několik samostatných vnímacích statických aparátů.
VII pár - lícní nerv, n. facialis, má jedno motorické jádro umístěné ve formacio reticularis partis dorsalis mostu pons. Nervová vlákna z něj vybíhající na své cestě tloušťkou můstku tvoří smyčku, která vyčnívá na kosočtverečné jámě v podobě colliculus facialis.
Střední nerv, n. intermedius, úzce spojený ve svém průběhu s lícním nervem, má dvě jádra:
1. Autonomní (sekreční), nucleus salivatorius superior (nadřazené slinné jádro), umístěné ve formacio reticularis mostu, dorzálně k jádru lícního nervu.
2. Citlivé, jádro trdctus solitdrii.
VI pár - abducens nerv, n. abdiicens, má jedno motorické jádro umístěné ve smyčce lícního nervu, proto tomuto jádru odpovídá colliculus facialis na povrchu kosočtverečné jamky
V pár - trojklaný nerv, n. trigeminus, má čtyři jádra:
1. Citlivý, nucleus sensorius principalis n. trigemini, se promítá do dorzolaterální části horní části mostu.
2. Jádro páteřního traktu, nucleus tractus spinalis n. n. trigemini, je pokračováním předchozího po celé délce prodloužené míchy až ke krční míše, kde se dostává do kontaktu se substantia gelatinosa hřbetních rohů.
3. Motor nucleus, nucleus motorius n. trigemini (žvýkací), umístěné mediálně k citlivému.
4. Jádro středního mozku, nucleus tractus mesencephalici n. n. trigemini, leží laterálně od akvaduktu. Představuje jádro proprioceptivní citlivosti pro žvýkací svaly a pro svaly oční bulvy.
Je možné, že toto jádro odráží samostatný vývoj první větev trojklaného nervu (n. ophthalmicus), u zvířat nazývaná n. ophthalmicus profundus a souvisí s orgánem vidění, což vysvětluje umístění jádra ve středním mozku.
Mezimozek
Střední mozek, mezimozek, se vyvíjí v procesu fylogeneze pod převažujícím vlivem zrakového receptoru, proto jeho nejdůležitější útvary souvisí s inervací oka. Zde se vytvořila centra sluchu, která spolu s centry zraku následně vyrostla v podobě čtyř tuberkul – quadrigeminu. S příchodem sluchových a vizuálních analyzátorů v kůře u vyšších zvířat a lidí přední mozek sluchová a zraková centra středního mozku sama upadla do podřízené pozice a stala se intermediální, subkortikální. S rozvojem předního mozku u vyšších savců a člověka začaly středním mozkem procházet dráhy spojující kůru terminálního provazce s míchou (mozkové stopky) (viz obr. 277, 278).
Výsledkem je, že lidský střední mozek obsahuje: 1) centra subkortikálního vidění a nervová jádra, která inervují oční svaly; 2) subkortikální sluchová centra; 3) všechny vzestupné a sestupné dráhy spojující mozkovou kůru s míchou a procházející středním mozkem; 4) svazky bílé hmoty spojující střední mozek s ostatními částmi centrálního nervového systému. V souladu s tím má střední mozek, který je nejmenší a nejjednodušeji strukturovanou částí mozku u lidí, dvě hlavní části: střechu, kde jsou umístěna subkortikální centra sluchu a zraku, a mozkové stopky, kde převážně procházejí dráhy.
1. Hřbetní část, střecha středního mozku nebo quadrigeminální ploténka, tectum mesencephali s. lamina quadrigemina(BNA) (viz obr. 278).
Je skryta pod zadním koncem corpus callosum a je rozdělena dvěma křížícími se rýhami - podélnou a příčnou - na čtyři bílé tuberkuly, uspořádané do párů.
Horní dva tuberkuly, colliculi superiores, jsou subkortikálními centry vidění; obě dolní, colliculi inferiores, jsou subkortikální sluchová centra. Tělo šišinky leží v ploché drážce mezi horními tuberkulami. Každý tuberkulum přechází v takzvanou rukojeť tuberkula, brachium colliculi, která jde laterálně, anteriorně a nahoru, k diencephalon. Rukojeť horního tuberkula, brdchium collculi superioris, jde pod polštář, pulvinar, optického tuberkula k laterálnímu geniculatu, corpus geniculatum laterale. Rukojeť dolního tuberkula, brdchium collculi inferioris, přecházející podél horního okraje trigonum lemnisci k sulcus lateralis mesencephali, mizí pod mediálním geniculatým tělem, corpus geniculatum mediale. Jmenovaná geniculate těla již patří do diencephalon.
2. Ventrální část, stopky mozku, pedunculi cerebri, obsahuje všechny cesty do předního mozku.
Mozkové stopky vypadají jako dva silné půlválcové bílé provazce, které se šikmo rozbíhají od okraje mostu a zanořují se do tloušťky mozkových hemisfér.
3. Dutina středního mozku, která je zbytkem primární dutiny měchýře středního mozku, má vzhled úzkého kanálu a je tzv. tekoucí voda,aqueductus cerebri(Sylvii). Je to úzký, ependymově lemovaný kanálek dlouhý 1,5-2,0 cm, spojující IV komoru s III. Dorzálně je akvadukt omezen střechou středního mozku a ventrálně tegmentem mozkových stopek.
Vnitřní struktura středního mozku. V příčném řezu středním mozkem se rozlišují tři hlavní části mozkového kmene: 1) střešní deska, lamina tecti, tvořený kvadrigeminálním; 2) pneumatika, tegmentum, představující horní část pedunculi cerebri; 3) ventrální část pedunculi cerebri nebo samotná mozková stopka, crus cerebri. Podle vývoje středního mozku pod vlivem zrakového receptoru (E.K. Sepp) obsahuje různá jádra související s inervací oka.
U nižších obratlovců slouží colliculus superior jako hlavní zakončení zrakového nervu a je zrakovým centrem. U savců a u lidí je při přesunu zrakových center do předního mozku zbývající spojení zrakového nervu s colliculus superior důležité pouze pro reflexy. Vlákna sluchové kličky (lemniscus lateralis) končí v jádře colliculus inferior a rovněž v mediálním geniculate. Střecha quadrigeminu má oboustranné spojení s míchou - trdctus spinotectdlis a trdctus tectobulbdris et tectospindlis. Posledně jmenované po dekusaci v tegmentu (dekusace Meynerta ve tvaru dorzální fontány) přecházejí do svalových jader v prodloužené míše a míchy. Jedná se o tzv. zrakově-zvukovou reflexní dráhu, která byla zmíněna při popisu míchy. Oblast kvadrigeminu lze tedy považovat za reflexní centrum pro různé druhy pohybů, které vznikají především pod vlivem zrakových a sluchových podnětů.
Sylvian akvadukt je obklopen centrální šedou hmotou, která svou funkcí souvisí s autonomním systémem. V něm, pod ventrální stěnou akvaduktu, v tegmentu mozkového stopky, jsou umístěna jádra dvou motorických mozkových nervů - n. oculomotorius(III pár) na úrovni colliculus superior a n. trochlearis(IV pár) na úrovni colliculus inferior. Jádro okulomotorického nervu se skládá z několika částí, které odpovídají inervaci několika svalů oční bulvy. Mediálně a za ním je další malé, také párové, vegetativní jádro, nucleus accessorius nebo Yakubovich jádro (Yakubovich to popsal v roce 1857, dříve než Westphal a Edinger, po kterém to bylo nesprávně nazýváno) a nepárové střední jádro. Yakubovičovo jádro a nepárové střední jádro inervují hladké svaly oka, m. ciliaris a m. sphincter pupillae. Tato část okulomotorického nervu patří do parasympatiku. Nad (orálním) jádrem okulomotorického nervu v tegmentu mozkového peduncle je jádro mediálního podélného fasciculu (Darkshevichovo jádro).
Laterálně od akvaduktu je jádro středního mozku n. trigeminus, nucleus tractus mesencephalici n. trigemini.
Mozkové stopky se dělí, jak již bylo zmíněno, na ventrální část, popř aktuální stopka mozku, crus cerebri, A pneumatika, tegmentum. Hranici mezi nimi tvoří černá látka, substantia nigra, která za svou barvu vděčí černému pigmentu obsaženému v jejích nervových buňkách – melaninu (obr. 284).

Substantia nigra se táhne po celé délce mozkové stopky od mostu po diencephalon; podle své funkce patří do extrapyramidového systému.
Crus cerebri, umístěný ventrálně k substantia nigra, obsahuje podélná nervová vlákna sestupující z mozkové kůry do všech pod nimi ležících částí centrálního nervového systému (tractus corticopontine, corticonuclearis, kortikospinální atd.). Tegmentum, nacházející se dorzálně k substantia nigra, obsahuje především různá vzestupná vlákna a také jádra šedé hmoty. Mezi nimi je nejvýznamnější nucleus ruber.
Tento protáhlý klobásovitý útvar se táhne v tegmentu mozkového stopky od subtuberkulární oblasti diencephalon k inferior colliculus, kde začíná důležitý sestupný trakt, tractus rubrospinalis, spojující červené jádro s předními rohy míšními. Tento svazek se po opuštění červeného jádra protíná s podobným svazkem opačné strany ve ventrální části sutury mediánu - ventrální dekusace tegmenta (Pstruh). Nucleus ruber je velmi důležité koordinační centrum extrapyramidového systému, propojené s jeho ostatními částmi. Vlákna do něj přecházejí z mozečku jako součást jeho horních stopek po jejich odříznutí pod kvadrigeminem, ventrálně z aqueductus cerebri a také z pallidum - to je nejnižší a nejstarší ze subkortikálních uzlin mozku. které jsou součástí extrapyramidového systému. Díky těmto spojením mozeček a extrapyramidový systém prostřednictvím červeného jádra a z něj vybíhajícího tractus rubrospinalis ovlivňují celý kosterní sval ve smyslu nevědomých automatických pohybů. Kromě sestupných podélných vláken obsahuje tegmentum vlákna vzestupná, tvořící pokračování mediální (lemniscus medialis) a laterální (lemniscus lateralis) kličky ve středním mozku. V rámci těchto smyček stoupají do velkého mozku všechny smyslové dráhy s výjimkou zrakové a čichové.
Pokračuje také tegmentum středního mozku retikulární formace, formatio reticularis, A fasciculus longitudindlis medialis. Ten druhý pochází z různá místa. Jedna z jeho částí začíná od vestibulárních jader, prochází na obou stranách po stranách střední čáry, přímo pod šedou hmotou dna Sylviova akvaduktu a IV komory, a skládá se ze vzestupných a sestupných vláken směřujících k jádrům hlavové nervy III, IV, VI a XI. Mediální podélný fasciculus je důležitá asociační dráha spojující mezi sebou různá jádra nervů očních svalů, která určuje kombinované pohyby očí, když se odchylují jedním nebo druhým směrem. S jeho funkcí souvisí i pohyby očí a hlavy, ke kterým dochází při stimulaci balančního aparátu.
Prosencephalon
Přední mozek, prosencephalon, se vyvíjí ve spojení s čichovým receptorem a zprvu (u vodních živočichů) jde o čistě čichový mozek, rhinencephalon. S přechodem zvířat z vodního prostředí do vzduchu se zvyšuje role čichového receptoru, protože s jeho pomocí určují látky obsažené ve vzduchu. chemické substance, signalizující zvíře o kořisti, nebezpečí a dalších životně důležitých přírodních jevech na velkou vzdálenost - vzdálený receptor.
Proto, a také díky vývoji a zdokonalování dalších analyzátorů, přední mozek u suchozemských zvířat výrazně roste a předčí ostatní části centrálního nervového systému a přeměňuje se z čichového mozku na orgán, který řídí celé chování zvířete. Podle dvou hlavních forem chování: 1) instinktivní chování založené na zkušenostech druhu (bez podmíněné reflexy) a 2) individuální chování založené na zkušenostech jednotlivce (podmíněné reflexy) - v předním mozku se vyvíjejí dvě skupiny center: 1) bazální neboli centrální uzly mozkových hemisfér („subkortex“); 2) mozková kůra. Tyto dvě skupiny center předního mozku přijímají všechny nervové vzruchy a sahají k nim všechny aferentní smyslové dráhy, které (až na výjimky) nejprve procházejí jedním společným centrem – zrakovým thalamem, neboli palácem, thalamem. Adaptace organismu na prostředí změnami metabolismu vedla v předním mozku ke vzniku vyšších center, která mají na starosti vegetativní procesy (subtalamická oblast, hypotalamus).
Ze dvou částí předního mozku, diencephalon, diencephalon a konečná telencephalon, kůra a subkortikální uzliny patří do telencephalon a zrakový talamus a subtalamická oblast patří do mezimozku. Podle E. K. Seppa (1959) se diencefalon nevyvinul v procesu fylogeneze z předního mozku v důsledku jeho rozdělení, ale společně se středním mozkem. To vysvětluje spojení zrakového receptoru nejen se středním mozkem, ale také s mezimozkem a jeho zrakovým thalamem, který proto dostal odpovídající název.
Diencephalon
Diencephalon, leží pod corpus callosum a fornix, po stranách srostlé s hemisférami telencephalon. Podle toho, co bylo řečeno výše o funkci a vývoji předního mozku, se v diencefalu rozlišují dvě hlavní části: 1) dorzální (fylogeneticky mladší) - thalamencefalon- centrum aferentních drah a 2) ventrální (fylogeneticky starší) - hypotalamu- nejvyšší vegetativní centrum. Dutina diencephalon je III komory.
Thalamencephalon
Thalamencephalon se zase skládá ze tří částí: thalamus – palác, epithalamus – suprathalamická oblast a metathalamus – transtalamická oblast (obr. 285).

A. Thalamus, thalamus, je velká párová akumulace šedé hmoty v laterálních stěnách diencefala po stranách třetí komory, mající vejčitý tvar, s předním koncem zašpičatělým ve formě tuberculum anterius a zadním koncem rozšířeným a zesíleným v forma polštáře, pulvinar. Rozdělení na přední konec a polštářek odpovídá funkčnímu rozdělení thalamu na centra aferentních drah (přední konec) a zrakové centrum (zadní). Hřbetní plocha je pokryta tenkou vrstvou bílé hmoty – stratum zonule. Ve svém laterálním řezu směřuje do dutiny postranní komory, oddělené od sousedního ocasatého jádra hraniční rýhou, sulcus termindlis, což je hranice mezi telencefalem, ke kterému patří ocasní jádro, a diencefalem, ke kterému optický tuberkul patří. Podél této rýhy se táhne pruh dřeně, stria termindlis.
Mediální povrch zrakového thalamu, pokrytý vrstvou centrální šedé hmoty, stojí svisle a směřuje k dutině třetí komory a tvoří její boční stěnu. Shora je od hřbetní plochy ohraničena bílým dřeňovým pruhem, stria medulldris thalami. Obě mediální plochy zrakových tuberosit jsou navzájem spojeny šedou komisurou - adhesio interthaldmica, ležící téměř uprostřed. Laterální plocha zrakového thalamu ohraničuje vnitřní burzu, capsula interna. Zrakový talamus se svou spodní plochou nachází nad mozkovou stopkou, splývající s její operkulem (obr. 286). Jak je vidět na řezech, šedá hmota zrakového thalamu je rozdělena bílými vrstvami, laminae medullares thalami, na samostatná jádra, pojmenovaná podle jejich topografie – přední, centrální, mediální, laterální a řada ventrálních.

Funkční význam vizuální thalamus je velmi velký. Přepínají se v ní aferentní dráhy: v jejím polštáři končí pulvinar, část vláken optického traktu (subkortikální centrum vidění), v předním jádře je svazek Vic d ́Azir, vycházející z corpora mamillaria a spojující tzv. zrakový tuberkul s čichovou sférou a nakonec všechny ostatní aferentní smyslové dráhy ze spodních částí centrálního nervového systému v jeho zbývajících jádrech, s lemniscus medialis končícím v laterálním jádře. Talamus je tedy subkortikálním centrem všech typy citlivosti Odtud jdou smyslové dráhy částečně do subkortikálních uzlin (díky kterým se thalamus opticus stává citlivým centrem extrapyramidového systému), částečně přímo do kůry (tractus thalamocorticalis).
B. Epithalamus. Strie medullares obou zrakových tuberosit směřují dozadu (kaudálně) a tvoří na obou stranách trojúhelníkové prodloužení, zvané trigonum habenulae. Z posledně jmenovaného pochází tzv. vodítko habenula, které se spolu se stejným vodítkem na opačné straně připojuje k tělu šišinky, corpus pineale (pinus - pine) s. epifýza (viz obr. 285). Před corpus pineale jsou obě vodítka svázána dohromady pomocí commissura habenuldrum. Samotné šišinka, trochu připomínající šišku (odtud pochází její název), svou stavbou a funkcí patří k žlázám s vnitřní sekrecí. Epifýza, vyčnívající zezadu do oblasti středního mozku, je umístěna v drážce mezi colliculi superior quadrigeminy a tvoří jakoby pátý tuberkulum.
V. Metathalamus. Za zrakovým thalamem jsou dvě malá vyvýšení – geniculate body, corpus geniculdtum laterale et mediate (viz obr. 278, 286).
Mediální geniculaté tělo menší velikosti, ale výraznější, leží před spodní rukojetí quadrigeminy pod pulvinárem optického thalamu, odděleno od něj jasnou rýhou. V ní končí vlákna sluchové kličky lemniscus lateralis, v důsledku čehož je spolu s dolními tuberkulami quadrigeminu subkortikálním centrem sluchu. Boční genikulovité tělo, větší, ve formě plochého tuberkula, se nachází na spodní boční straně pulvinaru. V něm z větší části končí laterální část optické dráhy (druhá část dráhy končí pulvinárem). Proto je laterální geniculate tělo spolu s pulvinarem a colliculus superior subkortikálním centrem vidění. Jádra obou geniculátních těl jsou spojena centrálními cestami s kortikálními konci odpovídajících analyzátorů.
Hypotalamus
Subtalamická oblast, hypothdlamus, v širokém slova smyslu sdružuje útvary umístěné ventrálně pod dnem třetí komory, před substantia perforata posterior, včetně subtuberkulární oblasti v užším smyslu, regio subthalamica. Podle embryonálního vývoje se hypotalamus dělí na dvě části: přední - vizuální, pars optica hypothalami, pod jehož názvem se kombinuje tuber cinereum s infundibulem a hypofýzou, dále chiasma opticum s tractus opticus a zadní - čichový: corpora mamillaria a regio subthalamica.
A. Tuber cinereum, šedý tuberkul, umístěný před corpora mamillaria, je nepárový dutý výběžek spodní stěny třetí komory, sestávající z tenké destičky šedé hmoty. Vrchol tuberkula je prodloužený do úzkého dutého trychtýře, infundibulum, na jehož slepém konci je mozkový přívěsek, hypofýza(glandula pituitaria), ležící v prohlubni sella turcica (její popis viz část „Orgány vnitřní sekrece“). Hlíza cinereum obsahuje jádra šedé hmoty, což jsou nejvyšší vegetativní centra ovlivňující zejména látkovou výměnu a termoregulaci.
B. Chiasma opticum, optické chiasma, leží před šedým tuberkulem, tvořeným optickým chiasmatem, nn. optika.
V. Corpora mamillaria, mastoidní tělíska, dvě malé bílé vyvýšeniny nepravidelně kulovitého tvaru, ležící symetricky po stranách střední čáry, před substantia perforata posterior. Pod povrchovou vrstvou bílé hmoty jsou uvnitř každého z těles dvě šedá jádra.
Tělíska mamillaria patří podle své funkce k subkortikálním čichovým centrům.
G. Regio subthalamica, podkožní oblast(v užším slova smyslu); Jedná se o malou oblast medully umístěnou pod optickým thalamem. V něm, laterálně od substantia nigra, leží oválné tělo, corpus subthalamicum Luysi, patřící do diencephalon. Lewisovo tělo je jedním z článků extrapyramidového systému; připisují se mu i vegetativní funkce.
III komory
III komora, ventriculus tertlus, se nachází těsně podél střední čáry a na přední části mozku vypadá jako úzká vertikální štěrbina. Laterální stěny třetí komory jsou tvořeny mediálními plochami zrakových tuberosit, mezi nimiž se téměř uprostřed rozprostírá adhesio interthalamica (viz obr. 285). Přední stěna Komoru tvoří zespodu tenká deska, lamina terminalis, a dále nahoru sloupy fornixu (columnae fornicis) s bílou přední komisurou ležící napříč, commissura cerebri anterior. Po stranách přední stěny komory omezují sloupy fornixu spolu s předními konci zrakových tuberositů interventrikulární foramina, foramina interventricularia (Monroi), spojující dutinu třetí komory s postranními komorami umístěnými v hemisférách telencephalon. Horní stěna Třetí komora, ležící pod fornixem a corpus callosum, je tela chorioidea ventriculi tertii (obr. 287), ta zahrnuje nedostatečně vyvinutou stěnu mozkového měchýře v podobě epiteliální destičky, lamina epithelialis a cévnatku srostlou s to. Po stranách střední čáry v tela chorioidea je vilózní plexus, plexus chorioideus ventriculi tertii. V oblasti zadní stěna komory jsou commissura habeniilarum et commissura cerebri posterior, mezi nimiž vystupuje do kaudální strany slepý výběžek komory, recessus pinealis. Ventrálně od commissura posterior ústí akvadukt do třetí komory trychtýřovitým otvorem. Spodní úzká stěna Třetí komora, ohraničená zevnitř od bočních stěn rýhami (sulci hypothalamici), ze strany mozkové báze odpovídá substantia perforata posterior, corpora marnillaria, tuber cinereum s chiasma opticum. V oblasti dna tvoří komorová dutina dvě prohlubně: recessus infundibuli, vyčnívající do šedého tuberkulu a do nálevky, a recessus opticus, ležící před chiasmem. Vnitřní povrch stěn třetí komory je pokryt ependymem.
Uvažované části mozku - kosočtverec (kromě mozečku), střední a střední jsou spojeny pod názvem mozkový kmen. Kliničtí lékaři někdy odkazují pouze na zadní mozek a střední mozek na mozkový kmen (E.K. Sepp, M.B. Zucker, E.V. Schmidt, 1950). Mozkový kmen, fylogeneticky starší útvar, se strukturou a funkcí výrazně liší od mladší části mozku – telencephalon.
telencephalon
Jak je zmíněno výše, telencephalon reprezentované dvěma hemisférami, hemispheria cerebri. Každá hemisféra obsahuje: plášť nebo plášť, pallium, čichový mozek, rhinencephalon a základní uzly. Zbytkem původních dutin obou váčků telencefala jsou postranní komory, ventriculi laterales. Přední mozek, ze kterého se uvolňuje telencefalon, se nejprve objevuje ve spojení s čichovým receptorem (čichový mozek), poté se stává řídícím orgánem chování zvířete a vznikají centra instinktivního chování na základě druhových reakcí (nepodmíněné reflexy). to - podkorové uzliny, a centra individuálního chování na základě individuální zkušenosti (podmíněné reflexy) - mozková kůra. Podle toho se v telencephalon rozlišují v pořadí historického vývoje: skupiny středisek:
1. Čichový mozek, rhinencephalon, je nejstarší a zároveň nejmenší částí, umístěnou ventrálně.
2. Bazální neboli centrální uzly hemisfér, „subkortex“, stará část telencephalon, paleencephalon, ukrytá v hlubinách.
3. Šedá hmota mozkové kůry, kůra, nejmladší část, neencephalon, a zároveň největší část, zakrývající zbytek jakoby pláštěm, odtud její název - plášť, neboli plášť, pallium.
Kromě dvou forem chování zaznamenaných u zvířat se u lidí objevuje třetí forma - kolektivní chování, založené na zkušenosti lidského kolektivu vytvořeného v tomto procesu. pracovní činnost lidská a lidská komunikace prostřednictvím řeči. Tato forma chování je spojena s vývojem nejmladších povrchových vrstev mozkové kůry, které tvoří materiální substrát tzv. druhého signálního (verbálního) systému reality (I. P. Pavlov).
Protože v procesu evoluce roste telencephalon nejrychleji a nejsilnější ze všech částí centrálního nervového systému, stává se největší částí mozku u lidí a nabývá vzhledu dvou objemných hemisfér - pravé a levé, hemispherium dextrum et sinistrum. V hloubce podélné trhliny mozku jsou obě hemisféry navzájem spojeny silnou vodorovnou deskou - corpus callosum, corpus callosum, který se skládá z nervových vláken probíhajících příčně z jedné hemisféry do druhé. V corpus callosum je přední konec ohnutý dolů neboli koleno, genu corporis calldsi, střední část, truncus corporis calldsi, a pak zadní konec, zesílený ve tvaru válečku, splenium corporis calldsi. Všechny tyto části jsou dobře patrné v podélném řezu mozkem mezi oběma hemisférami (viz obr. 282). Rod corpus callosum, zakřivený dolů, se zostřuje a tvoří zobák, rostrum corporis callosi, který přechází v tenkou desku lamina rostralis, která zase pokračuje v lamina terminalis.
Pod corpus callosum je tzv trezor, fornix(viz obr. 282; obr. 288), představující dvě obloukové bílé šňůry, které ve své střední části, corpus fornicis, jsou vzájemně propojeny a rozbíhají se vpředu a vzadu a tvoří obloukové sloupy vpředu, Сolumnae fornicis, vzadu jsou nohy oblouku, crura fornicis. Crura fornicis směřující dozadu sestupuje do dolních rohů postranních komor a přechází tam do fimbrie hippocampi. Mezi crura fornicis, pod splenium corporis callosi, se táhnou příčné svazky nervových vláken, které tvoří commissura fornicis. Přední konce fornixu, columnae fornicis, pokračují dolů k základně mozku, kde končí v corpora mamillares, procházející šedou hmotou hypotalamu. Columnae fornicis je ohraničeno za nimi ležícími interventrikulárními foraminami, spojujícími třetí komoru s postranními komorami. Před sloupy oblouku je přední komisura, commissura anterior, která vypadá jako bílá příčná tyč složená z nervových vláken. Mezi přední částí fornixu a genu corporis callosi je protažena tenká vertikální destička mozkové tkáně - průhledná přepážka septum pellucidum, v jejíž tloušťce je malá štěrbinovitá dutina cavum septi pellucidi (viz obr. 282, 293).
Rýže. 288. Fornix, hippocampus a přední komisura. 1 - corpus fornicis; 2 - cms fornicis; 3 - fimbria hippocampi; 4, 5 - columna fornicis; 6 - corpus mamillare; 7 - fasciculus thalamomamillaris (Vicq - d "Azyr); 8, 9 - comissura anterior; 10 - splenium corporis callosi; 11 - trigonum collaterale; 12 - calcar avis; 13 - polus occipitalis; 14 - 115 polus temporalis temporalis; 16 - hipokampus
Pro usnadnění studia začneme popis částí v opačném pořadí. historický vývoj, tedy z pláště, který zakrývá zbývající části.
pallium
V každé hemisféře lze rozlišit tři povrchy: superolaterální, mediální a dolní a tři okraje: horní, dolní a mediální, tři konce nebo póly: přední pól, polus frontalis, zadní, polus occipitalis a pak polus temporalis, odpovídající výběžek spodní plochy a na něm oddělen fossa, fossa lateralis cerebri (Sylvii).
Povrch polokoule (plášť), je tvořena stejnoměrnou vrstvou šedé hmoty o tloušťce 1,3-4,5 mm, obsahující nervové buňky. Tato vrstva, nazývaná také mozková kůra, cortex cerebri, vypadá jako složený, díky čemuž má povrch pláště vysoce komplexní kresba, skládající se ze střídání různé směry drážky a hřebeny mezi nimi, nazývané konvoluce *, gyri. Velikost a tvar rýh podléhá výrazným individuálním výkyvům, v jejichž důsledku nejen mozek odlišní lidé, ale ani hemisféry téhož jedince nejsou ve vzoru rýh zcela podobné (viz klasifikace rýh).
* (Celková plocha kůry dospělého člověka je asi 220 000 mm 2 (Geneberg), přičemž 2/3 leží hluboko mezi gyri a pouze 1/3 leží na povrchu.)
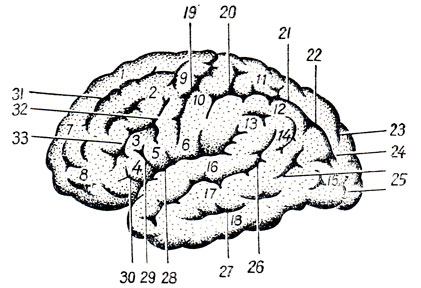
Hluboké, trvalé drážky se používají k rozdělení každé hemisféry do velkých oblastí nazývaných laloky. lobi; ty druhé se zase dělí na lalůčky a konvoluce. V každé hemisféře je pět laloků: čelní ( lobus frontalis), parietální ( lobus parietalis), dočasný ( lobus temporalis), týlní ( lobus occipitalis) a lalůček skrytý na dně laterální (Sylviánské) trhliny, tzv. ostrůvek ( ostrov).
Superolaterální povrch hemisféra je ohraničena na laloky třemi rýhami (viz obr. 274; obr. 289): laterální, střední a horní konec parietookcipitálního sulcus, který je na mediální straně hemisféry a tvoří na ní zářez. horní okraj. Laterální sulcus, sulcus cerebri lateralis(Sylvii), začíná na bazálním povrchu hemisféry od Sylvian fossa a pak přechází na superolaterální povrch, jde dozadu a mírně nahoru. Končí přibližně na hranici střední a zadní třetiny superolaterální povrch hemisféry. Vpředu laterální sulcus odcházejí z něj dvě malé větve: jedna, ramus ascendens, a druhá, ramus anterior, směřující k čelnímu laloku.

Centrální nebo Rolandová, sulcus, sulcus centralis(Rolandi), začíná na horním okraji polokoule, poněkud za jejím středem, a jde dopředu a dolů. Dolní konec centrálního sulku nedosahuje laterálního sulku. Část hemisféry nacházející se před centrálním sulkem patří k čelnímu laloku; část povrchu mozku ležící za centrálním sulkem tvoří parietální lalok, který je pomocí zadní části laterálního sulku vymezen od spodního temporálního laloku. Zadní hranice parietálního laloku je koncem výše uvedeného parietookcipitální sulcus, sulcus parietooccipitalis, nacházející se na mediální ploše hemisféry, ale tato hranice je neúplná, protože jmenovaná rýha nezasahuje daleko k superolaterální ploše, v důsledku čehož parietální lalok přímo přechází v okcipitální lalok. Tento druhý také nemá ostrou hranici, která by ho oddělovala od předního spánkového laloku. V důsledku toho je hranice mezi právě zmíněnými laloky nakreslena uměle pomocí čáry probíhající od parietookcipitálního sulcus ke spodnímu okraji hemisféry.
Každý lalok se skládá z řady konvolucí, někdy nazývaných laloky, které jsou ohraničeny drážkami na povrchu mozku.
Čelní lalok. V zadní části vnějšího povrchu tohoto laloku je sulcus precentralis téměř v paralelním směru k sulcus centralis. V podélném směru z něj vybíhají dvě drážky: sulcus frontalis superior et sulcus frontalis inferior. Tím čelní lalok je rozdělena na čtyři závity - jeden vertikální a tři horizontální. vertikální gyrus, gyrus precentralis, se nachází mezi sulcus centralis a sulcus precentralis.
Horizontální gyri čelního laloku následující: 1) přední frontální, gyrus frontalis superior, který jde nad sulcus frontalis superior, rovnoběžně s horním okrajem hemisféry, dosahuje jeho mediální plochy; 2) střední frontální gyrus, gyrus frontalis medius, táhne se mezi horní a dolní frontální sulci a 3) dolní frontální gyrus, gyrus frontalis inferior, uložený mezi sulcus frontalis inferior a laterální sulcus. Větve laterálního sulku, zasahující do gyru frontalis inferior, jej rozdělují na tři části: pars opercularis, ležící mezi dolním koncem sulcus precentralis a ramus ascendens sulci lateralis, pars triangularis, který se nachází mezi oběma větvemi laterálního sulku a konečně pars orbitalis, umístěný vpředu od ramus anterior sulci lateralis.
Parietální lalok. Na něm je umístěna přibližně rovnoběžně se středovou drážkou sulcus postcentralis, obvykle splývající s sulcus intraparietalis, jdoucí v horizontálním směru. V závislosti na umístění těchto rýh je temenní lalok rozdělen na tři gyri, z nichž jeden je vertikální a další dva jsou horizontální. vertikální gyrus, gyrus postcentralis, jde za sulcus centralis ve stejném směru jako gyrus precentralis, oddělený od něj centrální rýhou. Nad sulcus intraparietalis je horní parietální gyrus neboli lobule, lobulus parietalis superior, která sahá až k mediálnímu povrchu hemisféry. Pod sulcus intraparietalis leží lobulus parietalis inferior, která směrem dozadu obchází konce laterální rýhy a sulcus temporalis superior a ztrácí se v oblasti týlní lalok. Část lobulus parietalis inferior, která jde kolem laterálního sulku, se nazývá gyrus supramargindlis; druhá část, která jde kolem sulcus temporalis superior, se nazývá gyrus anguldris.
Temporální lalok. Boční plocha tohoto laloku má tři podélné gyry, vzájemně ohraničené sulcus temporalis superior A sulcus temporalis inferior. horní z konvolucí, gyrus temporalis superior, umístěný mezi laterální rýhou a sulcus temporalis superior. Jeho horní plocha, ukrytá v hloubce laterálního sulku, nese 2-3 krátké konvoluce zvané gyri tempordles transversi (Heschlovy konvoluce). se táhne mezi horními a dolními temporálními drážkami gyrus temporalis medius. Pod ním, oddělujícím se od něj, prochází sulcus temporalis inferior gyrus temporalis inferior, který je oddělen spodním okrajem od gyrus occipitotemporalis lateralis ležícího na spodní ploše.
Týlní lalok. Drážky na boční ploše tohoto laloku jsou proměnlivé a nekonzistentní. Z nich se rozlišuje příčná sulcus occipitalis transversus, obvykle navazující na konec sulcus intraparietalis.
Ostrůvek, ostrov. Chcete-li vidět tento lalůček, je nutné se oddálit nebo odstranit okraje boční drážky visící nad ním. Tyto okraje, patřící k čelnímu, parietálnímu a spánkovému laloku, se nazývají tegmentum, operculum. Ostrov má tvar trojúhelníku, jehož vrchol směřuje dopředu a dolů. Před, nad a za ostrůvkem je od sousedních částí ohraničen hlubokou drážkou, sulcus circleis. Povrch insula je pokryt krátkými záhyby.
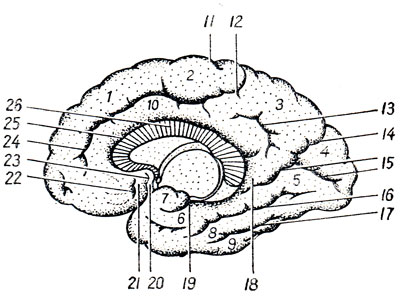
Spodní povrch polokoule(viz obr. 276) v té její části, která leží před Sylvian fossa, patří k čelnímu laloku. Zde probíhá paralelně s mediálním okrajem hemisféry sulcus olfactorius, ve kterém leží bulbus et tractus olfactorius. Mezi tímto sulkem a mediálním okrajem hemisféry se rozprostírá rovný gyrus, gyrus rectus, což je pokračování gyru frontalis superior. Laterálně od sulcus olfactorius na spodní ploše je několik nepravidelných rýh, sulci orbitales, omezující gyri orbitales, který lze považovat za pokračování středního a dolního frontálního gyru. Zadní část bazálního povrchu hemisféry je tvořena spodním povrchem temporálního a týlní lalok y, které zde nemají žádné určité hranice. V této oblasti jsou viditelné dvě drážky: sulcus occipitotemporalis, přecházející ve směru od okcipitálního pólu k časovému a omezujícímu gyrus occipitotemporalis lateralis a běží paralelně s ním sulcus kolaterály(jeho předním pokračováním je sulcus rhindlis). Mezi nimi se nachází gyrus occipitotemporalis medialis. Mediálně k sulcus collateralis jsou dva gyri: mezi zadní částí této rýhy a sulcus calcarinus leží gyrus lingualis; mezi přední částí této rýhy a sulcus rhinalis na jedné straně a hlubokým sulcus hippocampi, obklopujícím mozkový kmen, na straně druhé leží gyrus parahippocampalis. Tento gyrus, přiléhající k mozkovému kmeni, se již nachází na mediálním povrchu hemisféry.
Mediální povrch hemisféry. Na této ploše (viz obr. 282) je rýha corpus callosum, sulcus corporis callosi, probíhající přímo nad corpus callosum a pokračující svým zadním koncem do hloubky sulcus hippocampi, který směřuje dopředu a dolů. Paralelně a nad touto drážkou probíhá podél mediálního povrchu hemisféry sulcus cinguli, který začíná vpředu pod zobákem corpus callosum, pak jde zpět a končí svým zadním koncem na horním okraji hemisféry. Prostor nacházející se mezi tímto okrajem hemisféry a sulcus cinguli patří k frontálnímu laloku, k gyrus frontalis superior. Malá oblast nad sulcus cinguli, ohraničená za zadním koncem sulcus cinguli a vpředu malou rýhou, sulcus paracentrdlis, se nazývá cirkumcentrální lobule. lobulus paracentralis, protože odpovídá mediální ploše horních konců obou centrálních gyroskopů, které zde do sebe přecházejí.
Za lobulus paracentralis je čtyřúhelníková plocha (tzv. precuneus, precuneus), ohraničený vpředu koncem sulcus cinguli, dole malým sulcus subparietdlis a vzadu hlubokým sulcus parietooccipitalis. Precuneus odkazuje na parietální lalok. Za precuneem leží ostře oddělená oblast kůry patřící okcipitálnímu laloku, klínu, cuneus, který je vpředu omezen sulcus parietooccipitalis, a vzadu sulcus calcarfnus, sbíhající se pod úhlem. Inferiorně a posteriorně je klín v kontaktu s gyrus lingualis. Gyrus cingulate se rozprostírá mezi sulcus cinguli a žlábkem corpus callosum, gyrus cinguli, která přes šíji, šíje, pokračuje do gyrus parahippocampalis, končí háčkem, uncus. Gyrus parahipocampal je na jedné straně omezen sulcus hippocampi, který obepíná mozkový kmen, a na druhé straně sulcus collateralis a jeho předním pokračováním, zvaným sulcus rhinalis. Isthmus - zúžené místo přechodu gyrus cingulate do parahippokampus, se nachází za splenium corporis callosi, na konci rýhy vzniklé srůstem sulcus parietooccipitalis se sulcus calcaneus. Gyrus cinguli, isthmus a gyrus parahippocampalis tvoří společně klenutý gyrus, gyrus fornicatus, který popisuje téměř úplný kruh, otevřený pouze dole a vpředu. Klenutý gyrus nesouvisí s žádným z plášťových laloků.
Po odtažení okraje sulcus hippocampi je vidět úzký zubatý šedý pruh, což je základní gyrus gyrus dentatus.
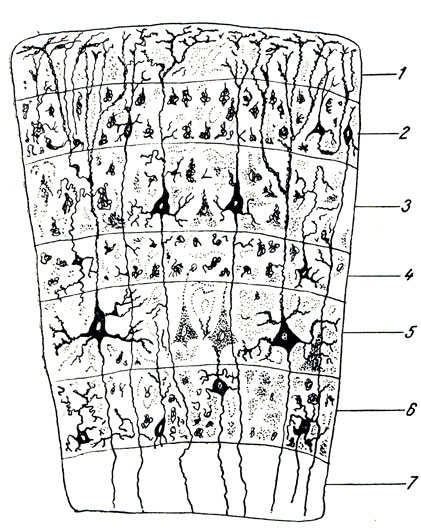
Stavba mozkové kůry. Kůra mozkových hemisfér se ve svém hlavním typu skládá ze šesti vrstev, lišících se od sebe především tvarem nervových buněk v nich obsažených (obr. 290): 1) molekulární vrstva leží přímo pod pia mater a obsahuje tzv. koncové větve procesů nervových buněk, propletené síťovitým způsobem; 2) vnější zrnitá vrstva, nazývaná tak, protože obsahuje četné malé buňky podobné zrnu; 3) pyramidální vrstva se skládá z malých a středně velkých pyramidálních nervových buněk; 4) vnitřní zrnitá vrstva je složena stejně jako vnější vrstva z malých buněk - zrn; 5) gangliová vrstva obsahuje velké pyramidální buňky (Betzovy buňky); 6) vrstva polymorfních buněk ohraničuje bílou hmotu. Z těchto 6 vrstev jsou spodní (5. a 6.) převážně začátkem eferentních drah; konkrétně vrstva 5 se skládá z Betzových pyramidálních buněk, jejichž axony tvoří pyramidový systém. Střední vrstvy (3. a 4.) jsou spojeny především s aferentními drahami a horní (1. a 2.) patří k asociativním drahám kůry. Šestivrstvý typ kůry je v různých oblastech modifikován, a to jak tloušťkou a uspořádáním vrstev, tak složením buněk (které je podrobně probráno v kurzu histologie).
Čichový mozek (rhinencephalon)
Čichový mozek, rhinencephaton(obr. 291, 292), je fylogeneticky nejstarší částí předního mozku, vznikající v souvislosti s čichovým receptorem, kdy se přední mozek ještě nestal orgánem zvířecího chování. Proto jsou všechny jeho součásti různými částmi čichového analyzátoru (koncept analyzátoru.

U ryb je orgánem čichu téměř celý přední mozek. S vývojem nové kůry, která je pozorována u savců a lidí, se vyvíjí nová část předního mozku (neencephalon) - plášť, pallium. Plášť ale také prochází dlouhou cestou vývoje a obsahuje tři části různých fylogenetických dat. Starší díly:
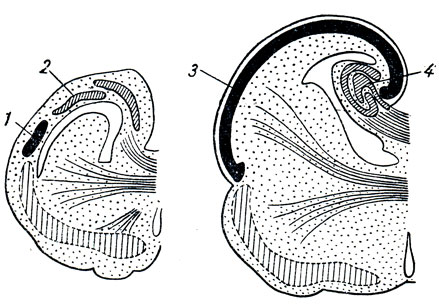
1. Archipalium, část spánkového laloku. Zpočátku se tento úsek nacházel na laterální ploše hemisféry, ale později se vlivem značně se zvětšujícího neopallia stočil do klobásovitého útvaru – hippocampu neboli Ammonova rohu a mediálně se posunul do dutiny postranní komora telencephalon v podobě výběžku jejího dolního rohu. Ammonův roh je pokryt starodávnou kůrou, archikortexem.
2. Paleopallium, malá oblast kůry na ventrálním povrchu čelního laloku, ležící blízko bulbus olfactorius a pokrytá starou kůrou, paleokortex.
3. Neopallium, nový plášť, v jehož kortexu, neokortexu, se objevila vyšší centra pachu - kortikální konce analyzátoru. Toto je uncus, který je součástí klenutého gyru.
V důsledku toho obsahuje lidský čichový mozek řadu útvarů různého původu, které lze topograficky rozdělit na dvě části. Periferním úsekem je čichový lalok, lobus olfactorius, který označuje řadu útvarů ležících na spodině mozku: 1) bulbus olfactorius; 2) tractus olfactorius; 3) trigonum olfactonum a 4) substantia perforata anterior. Centrální částí jsou konvoluce mozku: 1) gyrus mořského koníka, hippocampus; 2) gyrus dentatus, gyrus dentatus; 3) klenutý gyrus, gyrus fornicatus, s přední částí umístěnou v blízkosti temporálního pólu - hák, uncus.
Postranní komory


V hemisférách, jako pozůstatek původních dutin obou váčků telencephalon, leží pod úrovní corpus callosum symetricky po stranách střední čáry dvě postranní komory, ventriculi laterdles, oddělený od superolaterálního povrchu hemisfér celou tloušťkou dřeně. Dutina každé postranní komory (viz obr. 294) odpovídá tvaru hemisféry: začíná v čelním laloku ve formě zakřivené dolů a laterální strany přední roh, cornu anterius, odtud se táhne oblastí temenního laloku tzv centrální část, pars centralis a poté se na úrovni zadního okraje corpus callosum stáčí dolů a jde dopředu v tloušťce spánkového laloku ve tvaru dolní roh, cornu inferius, kde končí. V místě, kde komorová dutina klesá, vydává výběžek dozadu do týlního laloku - zadní roh, cornu posterius.
Mediální stěnu předního rohu tvoří septum pellucidum, které odděluje přední roh od stejného rohu druhé hemisféry (viz obr. 293). Boční stěnu a částečně i spodek předního rohu zaujímá šedá eminence, hlavice caudate nucleus, caput nuclei caudati a horní stěnu tvoří vlákna corpus callosum. Střecha centrální, nejužší části postranní komory je rovněž tvořena vlákny corpus callosum, zatímco dno tvoří pokračování caudatus nucleus, corpus nuclei caudati a část horní plochy zrakového thalamu. Hřbetní roh je obklopen vrstvou bílých nervových vláken pocházejících z corpus callosum, tzv. tapetum (koberec); na jeho mediální stěně je patrný hřeben - ptačí ostruha, calcar avis, tvořená prohlubní ze strany sulcus calcarinus, umístěnou na mediální ploše hemisféry. Superolaterální stěnu dolního rohu tvoří tapetum, které je pokračováním stejného útvaru obklopujícího zadní roh. Na mediální straně na horní stěně je ztenčená část nucleus caudate, ohnutá směrem dolů a vpředu - cauda nuclei caudati.
Podél mediální stěny dolního rohu je po celé jeho délce bílá elevace - Ammonův roh, cornu Ammonis, nebo mořský koník, hippocampus, který je tvořen v důsledku deprese z sulcus hippocampi hluboce se zařezávající do vnější strany. Přední konec hippocampu je rozdělen rýhami na několik malých tuberkul. Podél mediálního okraje amonského rohu se nachází tzv. třásněnka, fimbria hippocampi, která představuje pokračování pediklu oblouku (crus fornicis). Ve spodní části spodního rohu je hřeben, eminentia collaterdlis, který pochází z prohlubně mimo stejnojmennou rýhu. Z mediální strany postranní komory vystupuje do její centrální části a dolního rohu pia mater, která v tomto místě tvoří plexus choroideus, plexus chorioideus ventricular lateralis. Plexus je pokryt epitelem, který představuje zbytek nevyvinuté mediální stěny komory. Plexus chorioideus ventriculi lateralis je boční okraj těla chorioidea ventriculi tertii.
Bazální neboli centrální uzliny nebo jádra, hemisféry
Kromě šedé kůry mozkové na povrchu polokoule se v její tloušťce nacházejí i nahromadění šedé hmoty, tzv. bazální, centrální nebo subkortikální, jádra a tvoří to, co se zkráceně nazývá „subkortex“. Na rozdíl od kůry, která má strukturu screenových center, subkortikální jádra mají strukturu jaderných center. Existují tři shluky subkortikálních jader: corpus striatum, claustrum a corpus amygdaloideum (obr. 296, 297).
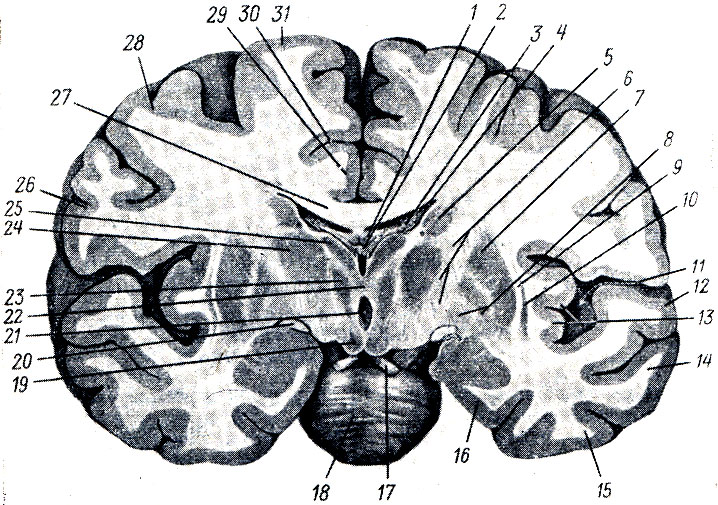
1. Corpus striatum, striatum, se skládá ze dvou částí, které nejsou od sebe zcela odděleny – nucleus caudatus a nucleus lentiformis.
A. Nucleus caudatus, caudate nucleus, leží nad a středově k nucleus lentiformis, oddělené od nucleus lentiformis vrstvou bílé hmoty nazývané vnitřní burza, capsula interna. Zesílená přední část caudate nucleus, jeho hlava, caput nuclei caudati, tvoří laterální stěnu předního rohu postranní komory, zatímco zadní ztenčený úsek nucleus caudate, corpus et cauda nuclei caudati, se táhne, jak máme my již vidět, zpět podél spodní části centrální části postranní komory; Kauda se ovíjí kolem horní stěny dolního rohu. Na mediální straně přiléhá nucleus caudatus k optickému thalamu, oddělený od něj pruhem bílé hmoty, stria terminalis. Zepředu a zespodu dosahuje hlava caudatus nucleus substantia perforata anterior, kde se spojuje s nucleus lentiformis (s jeho částí zvanou put amen). Kromě tohoto širokého spojení obou jader na ventrální straně existují také tenké proužky šedé hmoty umístěné proložené bílými svazky vnitřního pouzdra. Sloužily jako důvod k názvu - striatum, corpus striatum (viz obr. 296).
B. Nucleus lentiformis, lentikulární jádro, leží laterálně od nucleus caudatus a zrakového thalamu, oddělena od nich capsula interna. Na vodorovném řezu hemisférou má mediální plocha lentikulárního jádra, přivrácená k vnitřní burze, tvar úhlu s vrcholem směřujícím do středu; přední strana úhlu je rovnoběžná s ocasním tělem a zadní strana je rovnoběžná s vizuálním thalamem. Boční plocha je mírně konvexní a směřuje k laterální straně hemisféry v oblasti insuly. Zepředu a ventrálně, jak již bylo naznačeno, splývá lentikulární jádro s hlavicí nucleus caudatus. Na frontálním úseku má lentikulární jádro tvar klínu, jehož vrchol směřuje k mediální straně a báze směřuje k laterální straně. Čočkovité jádro je rozděleno na tři segmenty dvěma rovnoběžnými bílými vrstvami, laminae medullares, z nichž ta laterální, tmavě šedá, se nazývá skořápka, putamen(pleva) a dva střední, lehčí, se souhrnně nazývají globus pallidus, globus pallidus(viz obr. 296).
Globus pallidus, který se liší již svým makroskopickým vzhledem, má také histologickou strukturu, která je odlišná od ostatních částí striata. Fylogeneticky je globus pallidus starší útvar (paleostriatum) než putamen a nucleus caudatus (neostriatum).
Vzhledem ke všem těmto znakům je globus pallidus v současnosti řazen do zvláštní morfologické jednotky zvané pallidum, přičemž označení striatum je vyhrazeno pouze pro putamen a nucleus caudatus. V důsledku toho ztrácí termín „lentikulární jádro“ svůj předchozí význam a lze jej používat pouze v čistě topografickém smyslu a místo předchozího názvu corpus striatum se caudatum a lentikulární jádro nazývají striopallidální systém. Striopallidální systém je hlavní částí extrapyramidového systému (viz níže) a navíc je nejvyšším regulačním centrem autonomních funkcí ve vztahu k termoregulaci a metabolismu sacharidů, dominujícím nad podobnými autonomními centry v hypotalamu.
2. Claustrum, plot, představuje tenkou desku šedé hmoty, která se nachází v oblasti insula, mezi ní a putamen (viz obr. 297). Od druhé je oddělena vrstvou bílé hmoty, capsula externa, a od ostrovní kůry podobnou vrstvou zvanou capsula extrema.
3. Corpus amygdaloideum, amygdala nebo epistriatum, se nachází pod putamenem na předním konci spánkového laloku. Corpus amygdaloideum zřejmě patří k subkortikálním čichovým centrům. Končí svazkem vláken vycházejících z čichového laloku a substantia perforata anterior, poznamenaného v popisu zrakového tuberkulu zvaného stria terminalis (viz obr. 297).
Bílá hmota hemisfér
Veškerý prostor mezi šedou hmotou mozkové kůry a pletacími uzly je obsazen bílá hmota. Skládá se z velké množství nervová vlákna probíhající v různých směrech a tvořící dráhy telencefala. Nervová vlákna lze rozdělit do tří systémů: 1) asociační, 2) komisurální a 3) projekční vlákna.
A. Asociační vlákna(obr. 298) spojují různé části kůry téže hemisféry. Dělí se na krátké a dlouhé. Krátká vlákna, fibrae arcudtae cerebri, spojují sousední závity ve formě obloukovitých svazků. Dlouhá asociační vlákna spojují oblasti kůry, které jsou od sebe více vzdálené. Existuje několik takových svazků vláken. Cingulum, pás, svazek vláken procházející gyrus fornicatus, spojuje různé části kůry gyrus cinguli jak mezi sebou, tak se sousedními záhyby mediální plochy hemisféry. Frontální lalok je připojen k dolnímu parietálnímu laloku, týlnímu laloku a zadní části spánkového laloku přes fasciculus longitudindlis superior. Spánkový a týlní lalok jsou navzájem spojeny prostřednictvím fasciculus longitudindlis inferior. Nakonec je orbitální plocha frontálního laloku spojena se spánkovým pólem tzv. uncinate fasciculus, fasciculus uncindtus.

B. Komisurální vlákna, které jsou součástí tzv. mozkových komisur, neboli komisur, spojují symetrické části obou hemisfér. Největší mozkovou komisurou je corpus callosum, corpus callosum, který spojuje části obou hemisfér patřících k neencefalu.
Dvě mozkové komisury, commisstlra anterior et commissura fornicis, rozměrově mnohem menší, patří do rhinencephalon a spojují se: commissura anterior - čichové laloky a oba parahipokampální gyri, commissura fornicis - ammonovy rohy.
V. Projekční vlákna Spojují mozkovou kůru částečně s thalamus et corpora geniculata, částečně s podložními částmi centrálního nervového systému až po míchu včetně. Některá z těchto vláken vedou vzruchy centripetálně, směrem ke kůře, jiná naopak centrifugálně.
Projekční vlákna v bílé hmotě polokoule blíže ke kůře tvoří tzv. corona radiata, a pak hlavní část sbíhají se do výše zmíněného vnitřního vaku. Vnitřní burza, cdpsula interna, jak bylo uvedeno, je vrstva bílé hmoty mezi nucleus lentiformis na jedné straně a nucleus caudate a optickým thalamem na straně druhé. Na přední části mozku vypadá vnitřní burza jako šikmý bílý pruh, který pokračuje do mozkové stopky. Ve vodorovném řezu se jeví ve formě úhlu otevřeného k laterální straně (viz obr. 297), v důsledku čehož se v capsula interna rozlišuje přední část, crus anterius capsulae internae - mezi nucleus caudate a přední polovina vnitřního povrchu nucleus lentiformis, zadní část, crus posterius, mezi očním tuberkulem a zadní polovinou lentiformního jádra, a koleno, genu capsulae internae, ležící v inflexním bodě mezi oběma částmi vnitřní burza. Projekční vlákna podle jejich délky lze rozdělit do následujících systémů, počínaje nejdelším:
1. Trdctus kortikospinální(pyramidalis), vede motorické volní impulsy do svalů trupu a končetin. Počínaje pyramidovými buňkami kortexu střední a horní části předního centrálního gyru a lobulus paracentralis, vlákna pyramidového traktu procházejí jako součást corona radiata a poté procházejí vnitřním pouzdrem a zabírají přední dva- třetiny jeho zadní části, přičemž vlákna pro horní končetinu jdou před vlákny pro dolní končetiny. Poté procházejí mozkovou stopkou, crus cerebri, a odtud mostem do prodloužené míchy.
2. Trdctus kortikonukleární, vedoucí cesty k motorickým jádrům mozkových nervů. Počínaje pyramidálními buňkami kůry spodní části předního centrálního gyru procházejí kolenem vnitřní burzy a mozkovou stopkou, poté vstupují do mostu a pohybují se na druhou stranu a končí v motorických jádrech. z opačné strany, tvořící dekusaci. Malá část vláken končí bez křížení.
Vzhledem k tomu, že všechna motorická vlákna jsou shromážděna v malém prostoru ve vnitřní burze (koleno a přední dvě třetiny její zadní nohy), pokud jsou v tomto místě poškozena, jednostranná paralýza (hemiplegie) opačné strany těla je dodržován.
3. Trdctus corticopontini, dráhy z mozkové kůry do pontinních jader. Pocházejí z kůry čelního laloku (tractus frontopontinus), týlního (tractus occipitopontinus), temporálního (tractus temporopontine) a parietálního (tractus parietopontinus). Jako pokračování těchto drah jdou vlákna ve středních cerebelárních stopkách z pontinních jader do cerebellum. Pomocí těchto drah má mozková kůra inhibiční a regulační účinek na činnost mozečku.
4. Fasciculi thalamocortical et corticothaldmici, vlákna z thalamu do kůry a zpět z kůry do thalamu. Z vláken vycházejících ze zrakového thalamu je třeba si povšimnout tzv. tegmentální dráhy, což je závěrečná část smyslové dráhy směřující do centra kožního smyslu v zadním centrálním gyru. Vlákna této dráhy vycházející z laterálního jádra optického thalamu procházejí zadní částí vnitřního pouzdra za pyramidální dráhou. Tomuto místu se říkalo „citlivé chiasma“, protože tudy procházejí další smyslové dráhy, jmenovitě Graziolův optický svazek, radiatio optica, jdoucí z corpus geniculatum laterale a pulvinar zrakového thalamu do zrakového centra v kortexu týlního laloku. , dále sluchový svazek, radiatio acustica , směřující z corpus geniculatum mediale a dolního tuberkula quadrigeminu do Heschlových konvolucí ve spánkovém laloku, kde se nachází centrum sluchu. Zrakové a sluchové svazky zaujímají nejzadnější pozici v zadní části vnitřního pouzdra.
Mozek nachází se v mozkové části lebky. Hmotnost mozku dospělého je asi 1400-1500 g. U novorozence v průměru asi 380 g. Růst mozku končí o 23-25 let.
Mozek se skládá z pěti částí: přední, střední, střední, zadní (pons a cerebellum) a prodloužené míchy.
Fylogeneticky se nejstarší část mozku skládá z: prodloužené míchy, mostu, středního mozku a dvoumozku. Odtud vychází 12 párů hlavových nervů. Tato část tvoří mozkový kmen. Mozkové hemisféry se staly evolučně později.
Medulla je pokračováním míchy. Provádí reflexní a vodivé funkce. Vnějšek je pokrytý bílou hmotou, která tvoří dráhy mozku. Centrální část tvoří šedá hmota, kterou představují jednotlivé shluky neuronů. Tyto shluky tvoří jádra parasympatického nervového systému. V prodloužená medulla Nachází se zde tato střediska:
Respirační; srdeční činnost; vazomotorické; nepodmíněné potravinové reflexy; ochranné reflexy (kašel, kýchání, mrkání, slzení); centra změny tonusu určitých svalových skupin a polohy těla.
zadní mozek skládá se z mostu a mozečku. Dráhy mostu spojují prodlouženou míchu s mozkovými hemisférami.
Most se objevuje pouze u savců. V zadní části mostu je retikulární formace, jádra V-VIII párů hlavových nervů. Mozečkové stopky vybíhají z mostu. Most obsahuje jádra a dráhy sluchového analyzátoru.
Mozeček hraje hlavní roli při udržování tělesné rovnováhy a koordinace pohybů. Všichni obratlovci mají mozeček, ale úroveň jeho vývoje závisí na prostředí a povaze prováděných pohybů. Cerebellum má dvě hemisféry a střední část, vermis. Cerebellum je pokryto kůrou šedé hmoty. Mezi šedou hmotou jsou pruhy bílé hmoty. Mozeček se nejaktivněji vyvíjí mezi 5. a 11. měsícem života člověka. Růst cerebellum končí o 6-7 let a dosahuje 125-150 g.
Střední mozek během procesu evoluce se změnil méně než ostatní oddělení. Jeho vývoj je spojen s vizuálními a sluchovými analyzátory. Střední mozek zahrnuje dvě mozkové stopky a střechu (dlahy čtyřklanného nervu). Nohami procházejí vzestupné a sestupné nervové dráhy. Vzestupné dráhy jdou do thalamu, sestupné dráhy do prodloužené míchy a míchy. V kvadrigeminální oblasti jsou subkortikální centra pohybů, ke kterým dochází pod vlivem zrakových (superior colliculi) a sluchových (inferior colliculi) podnětů.
Diencephalon zahrnuje: zrakové pahorky (thalamus), supratuberkulární oblast (epitalamus), subtuberkulární oblast (hypotalamus) a genikulovitá těla. Obsahuje retikulární formaci - síť neuronů a nervových vláken, která ovlivňuje činnost různých částí centrálního nervového systému.
Thalamus je zodpovědný za všechny typy citlivosti (kromě čichové) a koordinuje mimiku, gesta a další projevy emocí. K thalamu přiléhá epifýza, což je žláza s vnitřní sekrecí. Do práce čichového analyzátoru se zapojují jádra epifýzy. Níže je další endokrinní žláza - hypofýza.
Hypotalamusřídí činnost autonomního nervového systému, regulaci metabolismu, homeostázu, spánek a bdění, endokrinní funkce organismu. Integruje nervové a humorální regulační mechanismy do společného neuroendokrinního systému. Hypotalamus tvoří s hypofýzou jeden komplex, ve kterém má řídící roli (řídí činnost předního laloku hypofýzy) a hypofýza má efektorovou roli. Hypotalamus vylučuje vazopresin a oxytocin, které vstupují do zadní hypofýzy a jsou odtud přenášeny krví.
Genikulovitá těla obsahují subkortikální centra zraku a sluchu.
Přední mozek se skládá z pravé a levé hemisféry spojených corpus callosum. Šedá hmota tvoří mozkovou kůru, bílá hmota- vodivé dráhy hemisfér. V bílé hmotě jsou rozptýlena jádra šedé hmoty (subkortikální struktury).
Mozková kůra se poprvé objevuje u plazů. U savců zaujímá většinu povrchu hemisfér a skládá se ze šesti vrstev buněk. Plocha kůry je asi 2-2,5 tisíc cm2. Tento povrch je spojen s přítomností velkého počtu drážek a záhybů.
Hluboké rýhy rozdělují hemisféru na čtyři laloky: čelní, parietální, temporální a okcipitální.
Spodní plocha hemisfér se nazývá základna mozku. Největšího rozvoje u člověka dosahují čelní laloky, oddělené od temenních laloků centrálním sulcusem. Jejich hmota tvoří asi 50 % hmoty mozku.
Oblasti mozkové kůry a jejich funkce:
Motorická zóna se nachází v předním centrálním gyru frontálního laloku;
Oblast muskulokutánní citlivosti se nachází v zadním centrálním gyru parietálního laloku;
Zraková oblast se nachází v okcipitálním laloku;
Sluchová zóna se nachází ve spánkovém laloku;
Centra čichu a chuti se nacházejí na vnitřních plochách spánkového a čelního laloku;
Asociační zóny kůry spojují její různé oblasti. Hrají si zásadní roli při tvorbě podmíněných reflexů.
Činnost všech lidských orgánů je řízena mozkovou kůrou. Jakýkoli míšní reflex se provádí za účasti mozkové kůry. Kůra poskytuje tělu vnější prostředí a je materiálním základem duševní činnosti člověka.
Funkční asymetrie je spojena s nepoměrem ve funkcích levé a pravé hemisféry. Pravá hemisféra je zodpovědná za imaginativní myšlení, levá za abstraktní myšlení. Když je poškozena levá hemisféra, je narušena řeč člověka.
Lidské tělo je velmi komplexní systém, který je řízen neuvěřitelně výkonným počítačem – mozkem. Vysílá signály všem orgánům a celému tělu, je zodpovědný za to, co cítíme, jak vnímáme svět a jak s ním komunikujeme. Uplynula staletí evoluce a lidé stále nebyli schopni plně pochopit celý mechanismus fungování mozku. Je hlavní složkou centrálního nervového systému.
Krátce o tom hlavním
Struktura lidského mozku je určena jeho funkcemi. Mozek je pokryt třemi typy membrán a celkem se skládá z 25 miliard neuronů, které se souhrnně nazývají šedá hmota. Ženský mozek váží o něco méně než mužský, ale to je způsobeno evolučním procesem, nikoli stupněm vývoje. V průměru jeho hmotnost dosahuje 2 % hmotnosti celého těla. Úroveň duševního vývoje nezávisí na velikosti mozku ani na velikosti těla.
Stavba mozkové kůry
V první řadě bychom si měli promluvit o kůře. Jeho tloušťka je 3 mm, pokrývá hlavní části mozku. Kůra má velmi složitou strukturu, skládá se ze šesti horizontálních vrstev, které se liší velikostí, hustotou atd. Má také vlastní specifické funkce. Některé z jeho laloků jsou zodpovědné za čich, dotyk, řeč atd.
Struktura mozku
Tento komplexní systém má několik oddělení, z nichž každé má své vlastní funkce. Rozlišují se následující hlavní oddělení:
- Poslední tvoří přibližně 80 % hmoty celého mozku.
- Střední - zde jsou thalamus a hypotalamus, které jsou zodpovědné za spojení člověka s okolím.
- Zadní - skládá se z mostu a mozečku, jejichž funkce spolu úzce souvisí.
- Prostřední je funkčně „nejchudší“.
- Oblongata - přímo spojená s míchou.
Kromě toho lze mozek také rozdělit do následujících částí:
- mozkové hemisféry;
- kmen;
- mozeček.
Konečný mozek

Toto oddělení je pravděpodobně nejsložitější jak strukturou, tak funkcí. Skládá se ze dvou částí: levé a pravé hemisféry, které jsou odděleny trhlinou. Uvnitř rýhy se zase nachází fornix a corpus callosum, které spojují hemisféry. Telencephalon je nejfunkčnější částí celého systému.
Levá hemisféra je zodpovědná za abstraktní myšlení a pravá hemisféra je zodpovědná za konkrétní myšlení. Kromě toho je telencephalon zodpovědný za řeč, emoční a analytické vnímání, získávání informací o životní prostředí a mnohem víc.
Diencephalon
Talamus je přímo zodpovědný za komunikaci s vnějším světem, reaguje na podněty a přenáší o nich informace do hemisfér. Hypotalamus reguluje autonomní funkce a interaguje s nervovým systémem. Pod ní se nachází hypofýza, jejíž funkce zahrnují regulaci spánku a bdění, metabolismus a kontrolu tělesné teploty.

zadní mozek
Funkce cerebellum a pons spolu úzce souvisí. Struktura mozku pomáhá pochopit, jak probíhá „komunikace“ mezi odděleními. Mozeček se nachází za mostem, který je částečně dirigentem. Mozeček se skládá ze šedé a bílé hmoty a je zodpovědný za koordinaci pohybů.
Střední mozek
Rozsah funkcí tohoto oddělení je malý, ale přesto velmi důležitý. Struktura lidské míchy je taková, že je přímo spojena s mozkem prostřednictvím středního provazce. Nese impulsy, které jsou způsobeny sluchovými a zrakovými podněty. Kromě toho je zodpovědný za tzv. skryté vidění a natáčení těla směrem k hluku. 
Medulla
Je přímo spojena s míchou. Struktura těchto dvou částí nervového systému má mnoho společného. Zde je bílá hmota, která slouží jako komunikační kanál procházející z míchy do mozku.
Mušle
Mozek je pokryt několika vrstvami membrán.
- Pia mater přímo interaguje s mozkem, pokrývá všechny konvoluce a sulci. Navíc se větví do samotného mozku a vyživuje ho.
- Arachnoidální membrána je nejtenčí, přichází do kontaktu s konvolucemi, ale nevyplňuje je.
- Tvrdá plena se skládá z velmi husté tkáně a je spojena s lebkou. Prostor mezi arachnoideou a dura mater je vyplněn serózní tekutinou.

- Mnoho propagandistů zdravý obraz V životě se říká, že alkohol zabíjí mozkové buňky, ale ve skutečnosti je to lež.
- Jednoho dne byl zastřelen maďarský voják a jeho přední lalok byl vážně poškozen. Přežil, ale do konce života nemohl spát.
- Existuje velmi rozšířená mylná představa, že lidé používají pouze malou část svého mozku. To je zásadně špatně. Náš mozek pracuje nepřetržitě a na plný výkon.
- Existuje také názor, že lidé s rozvinutější kreativní stránkou mají rozvinutější pravá hemisféra a lidé s analytickým myšlením mají rozvinutější levici. To není tak úplně pravda, ve skutečnosti jsou vyvíjeny stejně, jen je člověk díky tomu aktivnější.
závěry
Struktura mozku je velmi složitá. Výše uvedená oddělení a funkce jsou samozřejmě pouze základními znalostmi o tomto systému. Ve skutečnosti je toho v mozku hodně více prvků a jejich spojení jsou mnohem složitější. Dlouhý proces evoluce, během kterého se měnily funkce mozku, jeho velikost a tvar, vedl k tomu, že se mozek stal nejvýkonnějším „počítačem“ na Zemi.
Žádný stroj nemá tak vážné schopnosti, žádné zařízení se nedokáže tak rychle vypořádat s přidělenými úkoly. Jde o tak složitý systém, že ani za století neuvěřitelného pokroku ve všech oblastech vědění lidé nebyli schopni plně studovat mozek a nikdo nemůže s jistotou říci, že se jim to někdy podaří.



