Lobes and convolutions of the cerebral hemispheres. Brain
The brain, with its surrounding membranes, is located in the cranial cavity. The upper ventral surface of the brain corresponds in shape to the inner concave surface of the cranial vault. The lower surface, the base of the brain, has a complex relief corresponding to the cranial fossae of the inner base of the skull.
The mass of the adult human brain ranges from 1100 to 2000. Over the course of 20 to 60 years, the mass and volume remain maximum and constant for each individual.
When examining a specimen of the brain, its three largest components are clearly visible. These are paired hemispheres big brain, cerebellum and brain stem.
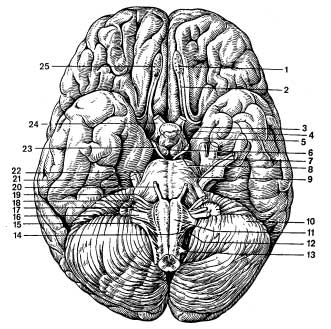
Figure A Base of the brain and exit of cranial nerve roots
HEMISPHERES OF THE BRAIN in an adult, this is the most highly developed, largest and functionally most important part of the central nervous system. The divisions of the hemispheres cover all other parts of the brain. Right and left hemisphere separated from each other by a deep longitudinal gap big brain, reaching the greater commissure of the brain, or corpus callosum.
In the posterior sections, the longitudinal fissure flows into transverse slot cerebrum, which separates the hemispheres from the cerebellum.
On the ventral, medial and lower surfaces of the cerebral hemispheres there are deep and shallow furrows. Deep grooves divide each of the hemispheres into lobes of the cerebrum. Small furrows separate from each other convolutions big brain. The lower surface, or base, of the brain is formed by the ventral surfaces of the cerebral hemispheres, the cerebellum and the ventral parts of the brain stem.
Adjacent to the posterior surface of the optic chiasm gray bump, the lower sections of which are elongated in the form of a tube gradually tapering towards the bottom - a funnel. At the lower end of the funnel there is a rounded formation - pituitary. Adjacent to the gray mound are two white spherical elevations - mastoid bodies.
Behind the optic tracts two longitudinal white ridges are visible - cerebral peduncles, between which there is a recess - interpeduncular fossa. Its bottom is formed posterior perforated substance. Even further away there is a wide transverse roller - bridge. The lateral sections of the pons continue into the cerebellum, forming it middle cerebellar peduncles.
Sections caudal to the bridge medulla oblongata presented medially located pyramids, separated from each other by the anterior medial fissure, and laterally - olives.
Inspection medial surface the cerebral hemispheres, some parts of the brain stem and the cerebellum become possible by making a midline incision along the longitudinal fissure of the cerebrum.
The extensive medial surface of the cerebral hemispheres hangs over the much smaller cerebellum and brain stem. On the medial surface of the hemispheres, as on other surfaces, grooves are visible that separate the gyri from each other.
Areas of the frontal, parietal and occipital lobes are separated from the corpus callosum by the groove of the same name.
The middle part of the corpus callosum is called trunk. Its anterior sections bend towards the bottom, forming knee corpus callosum. Even further down, the corpus callosum becomes thinner and passes into the beak of the corpus callosum. The posterior sections of the corpus callosum, in its middle part, a thin white plate is separated, called body of the arch. Gradually separating from the corpus callosum and forming an arcuate bend forward and downward, the body continues into the column of the vault, which ends with the mastoid body, and from behind - into the crura of the vault. A bundle of nerve fibers runs transversely between the pillars of the fornix, visible on the cut in the form of a white oval - this anterior commissure brain Like the transverse fibers of the corpus callosum, they connect the cerebral hemispheres with each other.
The columns of the fornix surround a thin plate of medulla - transparent partition.
All of the listed brain formations belong to the telencephalon.
The structures located below, with the exception of the cerebellum, belong to the brain stem (intermediate, middle, posterior parts of the brain and medulla oblongata).
The most anterior parts of the brain stem are formed by the visual tuberosities, which are located inferior to the body of the fornix and the corpus callosum and behind the columns of the fornix.
In a midline section of the brain, only the medial surface of the posterior thalamus (optic thalamus) is visible.
In the posterosuperior parts of the optic hillocks there is pineal gland, the anterior inferior sections of which are fused with a thin transversely running cord (posterior commissure).
The visual tuberosities and the formations located next to them, described above, belong to diencephalon.
Adjacent to the posterior surface of the optic thalamus are formations related to midbrain.
MEDULLA is a direct continuation of the spinal cord. The border between the medulla oblongata and the spinal cord corresponds to the level of the edges of the foramen magnum. The upper border of the medulla oblongata on the ventral surface runs along the posterior edge of the pons.
The anterior sections of the medulla oblongata become somewhat thicker compared to the posterior sections, and this section of the brain takes on the shape of a truncated cone. The grooves of the medulla oblongata are a continuation of the grooves of the spinal cord and have the same names. On both sides of the anterior median fissure on the ventral surface of the medulla oblongata there are convex pyramids gradually tapering towards the bottom.
Lateral to the pyramid on both sides there are oval elevations - olives.
In the lower part, on the dorsal surface of the medulla oblongata, there is a posterior median groove, on the sides of which the thin and wedge-shaped bundles of the posterior cords of the spinal cord end with thickenings. In these thickenings are located the nuclei of these bundles, from which the fibers depart, forming the medial loop. The medial loop at the level of the medulla oblongata forms a decussation. The bundles of this precross are located dorsal to the pyramids, in the interolive layer. The fibers of the medial bundle also pass here. Lateral to the olive, thin roots of the glossopharyngeal, vagus and accessory nerves emerge from the posterior lateral sulcus, the nuclei of which lie in the dorsolateral parts of the medulla oblongata.
The gray matter of the medulla oblongata is represented in the ventral sections by clusters of neurons that form the inferior olivary nuclei. Dorsal to the pyramids along the entire medulla oblongata there is a reticular formation, which is represented by an interweaving of nerve fibers and nerve cells lying between them.
At the level of the medulla oblongata there are vital centers such as respiratory and circulatory centers.
BRIDGE at the base of the brain stem it has the appearance of a transversely located white ridge, which has the appearance of a transversely located white ridge, which in the caudal region borders on the pyramids and olives of the medulla oblongata, and in the cranial region - on the cerebral peduncles.
The continuation of the bridge in the lateral direction forms middle cerebellar peduncle.
The dorsal surface of the pons is covered by the cerebellum and is not visible from the outside.
On a cross-section of the bridge in the central sections, you can see a thick bundle of fibers running transversely, related to the conduction path of the auditory analyzer and forming a trapezoidal body.
In the lower parts of the pons, there are noticeable accumulations of gray matter, called the nuclei of the pons proper, which act as intermediaries in the connections between the cerebral cortex and the cerebellar hemispheres.
The fibers lie in the dorsal part of the bridge medial lemniscus, coming from the medulla oblongata. above which is located reticular formation of the bridge. The fibers of the auditory lemniscus pass more laterally.
CEREBELLUM constitutes a larger part of the hindbrain than the pons, which fills most of the posterior cranial fossa.
In the cerebellum, there are superior and inferior surfaces, the boundaries between which are the anterior and posterior edges.
The upper surface of the cerebellum on the whole brain is covered by the occipital lobes of the cerebral hemispheres and is separated from them by the deep transverse fissure of the cerebrum. In the cerebellum there is an unpaired middle part - worm, two hemispheres. The worm is divided into small convolutions by transverse grooves, which give it some resemblance to an annelid worm. Both surfaces of the hemispheres and the vermis are cut by many transverse parallel small grooves, between which there are long and narrow convolutions of the cerebellum. A group of gyri separated by deeper grooves form the cerebellar lobules. The cerebellar hemispheres and the vermis consist of white matter located internally and a thin layer of gray matter of the cerebellar cortex bordering the white matter at the periphery. The cerebellar cortex is composed of three layers of nerve cells. On a sagittal section, the white matter of the cerebellum is represented by three layers of nerve cells and has the appearance of a branched tree.
In the thickness of the white matter, separate paired clusters of nerve cells are found, which form jagged, cork-shaped, spherical cerebellar nuclei and nuclei tent.
In the brain stem, the next section after the pons, small but functionally important, is the isthmus of the rhombencephalon, consisting of the superior cerebellar peduncles, the superior medullary velum and the triangular lemniscus, in which the fibers of the lateral (auditory) lemniscus pass.
MIDDLE BRAIN consists of the dorsal part of the roof of the midbrain and the ventral - cerebral peduncle, which are delimited by a cavity - the aqueduct of the brain. The lower border of the midbrain on its ventral surface is the anterior edge of the pons, the superior optic tract and the level of the mammillary bodies. On a brain specimen, the lamina tetracholomius, or the roof of the midbrain, can be seen only after the cerebral hemispheres have been removed.
At the base of the brain, the second part of the midbrain is clearly visible in the form of two thick white diverging bundles going into the tissue of the cerebral hemispheres - this is cerebral peduncles. The depression between the right and left cerebral peduncles is called interpeduncular fossa, the roots of the oculomotor nerves emerge from it. In front of the nucleus of the oculomotor nerve lies the nucleus of the medial longitudinal fasciculus. The largest nucleus of the midbrain is the red nucleus - one of the central coordination nuclei of the extrapyramidal system. Next to the aqueduct lies the reticular form of the midbrain.
The cross section clearly shows cranial substance, which divides the cerebral peduncle into two sections: dorsal - tegmentum of the midbrain and ventral - base of the cerebral peduncle. The midbrain nuclei are located in the tegmentum and ascending pathways pass through. The ventral parts of the cerebral peduncles consist entirely of white matter; descending pathways pass here.
The functional significance of the midbrain is: that the subcortical centers of hearing and vision are located here; nuclei of the head nerves that provide innervation to the striated and smooth muscles of the eyeball: nuclei belonging to the extrapyramidal system, which provides contraction of the body muscles during automatic movements. Descending (motor) and ascending (sensory) pathways follow through the midbrain. The midbrain area is also the location of the autonomic centers (central gray matter) and the reticular formation.
DENAMEBRAIN represented by the following departments:
- the area of the visual hillocks (thalamic area), which is located in its dorsal areas;
- the hypothalamus (subthalamic region), which makes up the ventral sections of the diencephalon;
- III ventricle, which has the form of a longitudinal (sagittal) fissure between the right and left visual tuberosities and connects through the interventricular foramen with the lateral ventricles.
The visual thalamus consists of gray matter, in which individual clusters of nerve cells (nuclei of the visual thalamus) are distinguished, separated by thin layers of white matter. Due to the fact that most of the sensitive pathways switch here, the visual thalamus is actually a subcortical sensory center, and its cushion is a subcortical visual center.
It is attached to the medial surface of the visual tuberosities using leashes. pineal body - pineal gland.
The hypothalamus makes up the ventral part of the diencephalon and is involved in the formation of the floor of the third ventricle. The hypothalamus includes gray bump with funnel And pituitary gland- iron internal secretion, optic tract, optic chiasm, mastoid bodies.
The hypothalamus is a continuation of the cerebral peduncles into the diencephalon. The gray matter of the subthalamic region is located in the form of nuclei capable of producing neurosecretion and transporting it to the pituitary gland, regulating the endocrine functioning of the latter.
Thus, the gray matter of the diencephalon consists of nuclei related to the subcortical centers of all types of sensitivity. In the area of the diencephalon there are the reticular formation, centers of the extrapyramidal system, vegetative centers that regulate all types of metabolism and neurosecretory nuclei.
White matter The diencephalon is represented by conducting pathways of ascending and descending directions, providing bilateral communication of the cerebral cortex with subcortical formations and centers of the spinal cord. In addition, the diencephalon includes two endocrine glands - the pituitary gland and the pineal gland, which, together with the corresponding nuclei of the hypothalamus and epithalamus, participate in the formation of the hypothalamic-hypophyseal and epithalamic-epiphyseal systems.
END BRAIN consists of two cerebral hemispheres, each of which is represented cloak, olfactory brain And basal ganglia. Cavity telencephalon are the lateral ventricles located in each hemisphere. The hemispheres of the cerebrum are separated from each other by the longitudinal fissure of the cerebrum and are connected by the corpus callosum, the anterior and posterior commissures and the commissure of the fornix. The corpus callosum consists of transverse fibers that continue in the lateral direction into the hemispheres, forming the radiation of the corpus callosum, connecting parts of the frontal and occipital lobes of the hemispheres with each other, bending in an arcuate manner and forming the anterior - frontal and posterior - occipital forceps. Adjacent to the posterior and middle parts of the corpus callosum below is the fornix of the brain, consisting of two arched cords fused in its middle part with the help of the anterior commissure of the brain.
CEREBRAL CORTEX formed by white and gray matter. There are 6 layers of nerve cells in the cortex, its different sections have different thicknesses (from 1.5 to 5.0 mm, on average 2-3 mm). Each of the hemispheres has three surfaces: the most convex upper lateral, flat, medial, facing the opposite hemisphere and having a complex relief corresponding to the internal base of the skull - the lower surface of the hemisphere or the base of the brain. The most prominent areas of the hemispheres are called the frontal, occipital, and temporal poles. The surface of the hemispheres is indented with deep cracks and grooves. The terrain located between them complicates the terrain - convolutions. The depth, length of the furrows, their shape and direction are very variable.
The fissures and grooves divide the hemispheres into frontal, parietal, temporal, occipital and insular lobes. The latter is not visible when viewing the surfaces of the hemispheres, because the insula is located at the bottom of the lateral sulcus and is covered by areas of other lobes.
On the superolateral surface of the hemisphere attracts attention lateral groove, which is the boundary between the frontal, parietal and temporal lobes and runs from the lower surface of the hemispheres back and up.
Another large furrow - central sulcus. It starts approximately from the middle of the upper edge of the hemispheres and follows down and somewhat forward, but does not reach the lateral sulcus. The central sulcus separates the frontal lobe from the parietal lobe. There is no pronounced border between the parietal and occipital lobes on the dorsolateral surface of the hemispheres.
Frontal lobe. In front of the central sulcus, the precentral sulcus stretches almost parallel to it, which gives rise to two parallel sulci running towards the frontal pole. These grooves divide the surface of the brain into those lying in front of the central sulcus precentral gyrus and horizontally running upper, middle And inferior frontal gyrus.
Parietal lobe. Posterior to the central sulcus and almost parallel to it runs postcentral sulcus, from which the longitudinal intraparietal sulcus. These two grooves divide the parietal lobe into postcentral gyrus, as well as on top And inferior parietal lobules.
Temporal lobe. The superior lateral surface of the temporal lobe is represented by two grooves running parallel lateral sulcus, which divide the surface of the brain into upper, middle And inferior gyrus.
The gray matter of the cerebral hemispheres is represented by the cortex and basal ganglia of the telencephalon. TO basal ganglia include the striatum, consisting of the caudate and lenticular nuclei; fence and amygdala. The layers of white matter between them form the outer and inner capsules, the latter being a thick layer of white matter consisting of the brain's pathways. The anterior and posterior legs and knee are distinguished in the internal capsule.
Striopallidar system represents the main part of the motor centers belonging to the extrapyramidal system. This is the center that controls automatic movements and regulates muscle tone. In addition, the striatum serves as a higher center that regulates the processes of heat regulation and carbohydrate metabolism. This center occupies a dominant position in relation to similar vegetative centers, which are located in the hypothalamic region.
The cerebral cortex is represented gray matter located on their periphery.
WHITE MATTER HEMISPHERES The cerebrum forms a white semi-oval center, which consists of a huge number of nerve fibers. All nerve fibers are represented by three systems of pathways in the telencephalon:
- associative;
- commissural;
- projection.
The functioning and interconnection of associative, commissural, as well as ascending and descending pathways ensures the existence of complex reflex arcs that allow the body to adapt to the constantly changing conditions of the internal and external Environment.
LATERAL VENTRICLES are located in the thickness of the white matter of the cerebral hemispheres. The cavity of the ventricles has a bizarre shape due to this. that the departments of each of them are located in all lobes of the hemisphere (with the exception of the insula). Average - central- part of the ventricle lies downward from the corpus callosum, in the parietal lobe of the hemisphere. From the central part, cavities called horns diverge into all lobes of the brain: front(frontal horn) - into the frontal lobe, lower(temporal horn) - into the temporal, rear- (occipital horn) - into the occipital lobe. The central part is connected to the III ventricle.
MEMBRANES OF THE BRAIN
The brain, like the spinal cord, is surrounded by three connective tissue sheets, or membranes, which are a continuation of the membranes of the spinal cord, each of which is separated from its neighbors by the intermeningeal space.
Dura mater of the brain. differs in structure from the similar membrane of the spinal cord. It is also the periosteum on the inner surface of the skull bones, with which it is loosely connected. In the area of the base of the skull, the shell gives off a number of processes that penetrate into the cracks and openings of the skull bones, which explains the greater strength of the attachment of the dura mater of the brain here. Moreover, in the places where the cranial nerves exit the cavity, the dura mater of the brain continues to surround the nerve for some extent, forming its sheath and penetrating together with the nerve through the opening to the outside.
On the inner surface of the dura mater, several processes are distinguished that penetrate the longitudinal fissure of the cerebrum and separate its hemispheres from each other. The posterior part of the falx fuses with another process of the membrane - the tentorium cerebellum, which separates the occipital lobes of the hemispheres from the cerebellum.
The continuation of the falx cerebri is falx cerebellum, penetrating from below between the cerebellar hemispheres. Another process surrounds the sella turcica on top, forming its diaphragm and protecting the pituitary gland from the pressure of the entire overlying mass of the brain.
In certain areas of the dura mater of the brain there are splits lined from the inside with endothelium - these are sinuses the dura mater of the brain through which venous blood flows. A special feature of the sinuses is the strength of the walls, which explains the impossibility of their collapse. In addition, the sinuses connect to the external veins of the head through emissary veins.
The arachnoid membrane of the brain is located medially from the dura mater and is separated from it by the subdural space.
The subarachnoid space of the brain in the region of the foramen magnum communicates with the subarachnoid space of the spinal cord.
In certain places, near the sinuses of the dura mater of the brain, the arachnoid membrane forms peculiar outgrowths - granulation of the arachnoid membrane. These outgrowths protrude into the sinuses of the dura mater. On the inner surface of the skull bones, at the location of the granulations, depressions and dimples are noted.
The generally accepted opinion is that granulation of the arachnoid membrane participates in ensuring the outflow of cerebrospinal fluid into the venous bed.
The pia (choroid) membrane is the innermost of the membranes of the brain. It consists of connective tissue that forms two layers (inner and outer), between which blood vessels lie. The membrane is fused to the outer surface of the brain and penetrates deeply into all its crevices and grooves. Blood vessels, leaving the choroid, go to the brain tissue, providing it with nutrition. In certain places, the choroid penetrates the cavities of the ventricles of the brain and forms choroid plexuses that project the cerebrospinal fluid.
Bibliography:
- Human anatomy R.P. Samusev Yu.M. Celine M.: Medicine 1995
- Human Physiology /ed. G. and Kositsky M.: Medicine 1985
In each hemisphere the following surfaces are distinguished:
convex superolateral surface adjacent to the inner surface of the bones of the arch skulls
bottom surface, the anterior and middle sections of which are located on the inner surface of the base of the skull, in the area of the anterior and middle cranial fossae, and the posterior ones - on tentorium cerebellum
medial surface, directed towards the longitudinal fissure of the brain .
These three surfaces of each hemisphere, passing one into another, form three edges. The superior edge separates the superolateral and medial surfaces. The inferolateral edge separates the superolateral surface from the inferior one. The inferomedial edge is located between the inferior and medial surfaces.
Each hemisphere consists of four lobes: frontal, parietal, temporal and occipital. In the cortex frontal lobes contains centers that regulate motor activity, as well as, probably, centers for planning and foresight. In the cortex of the parietal lobes, located behind the frontal lobes, there are zones of bodily sensations, including touch and joint-muscular sensation. Adjacent to the parietal lobe is the temporal lobe, in which the primary auditory cortex, as well as the centers of speech and other higher functions, are located. The posterior parts of the brain are occupied by the occipital lobe, located above the cerebellum; its cortex contains areas of visual sensation.
CORTEX covers the surface cerebral hemispheres with its numerous grooves and convolutions, due to which the area of the cortex increases significantly. There are association zones of the cortex, as well as sensory and motor cortex - areas in which neutrons are concentrated, innervating various parts of the body.
Areas of the cortex not directly associated with the regulation of movements or the analysis of sensory information are called the associative cortex. In these specialized zones, associative connections are formed between different areas and parts of the brain and the information coming from them is integrated. The association cortex supports complex functions such as learning, memory, language, and thinking.
52. Telencephalon. Hemispheres of the brain. Lobes, furrows, convolutions.
TO 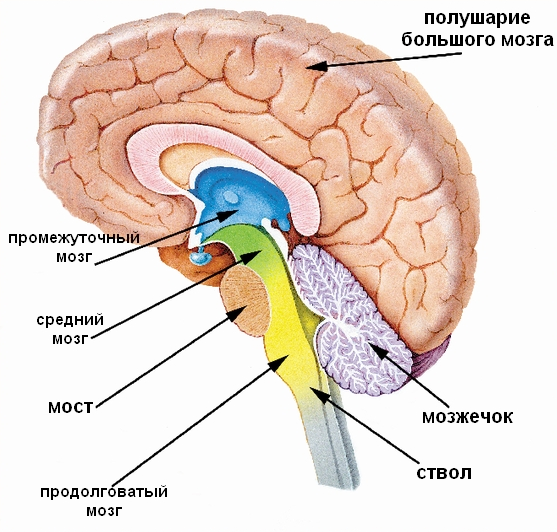 telencephalon- the most anterior part of the brain. Consists of two cerebral hemispheres(covered by the cortex), corpus callosum, striatum and olfactory brain. It is the largest section of the brain. It is also the most developed structure, covering all parts of the brain.
telencephalon- the most anterior part of the brain. Consists of two cerebral hemispheres(covered by the cortex), corpus callosum, striatum and olfactory brain. It is the largest section of the brain. It is also the most developed structure, covering all parts of the brain.
The cerebrum consists of two hemispheres, each of which is represented by the mantle, the olfactory brain and the basal ganglia. The cavity of the telencephalon is the lateral ventricles, located in each hemisphere. The hemispheres of the cerebrum are separated from each other by the longitudinal fissure of the cerebrum and are connected by the corpus callosum, the anterior and posterior commissures and the commissure of the fornix.
The surface of the cerebral cortex consists of folds - convolutions. They are separated by grooves; the shallow ones are called cerebral sulci, the deep ones are called cerebral fissures.
The telencephalon is divided into main lobes, which differ in both location and function:
Frontal lobe; - parietal lobe; - occipital lobe; - temporal lobe; - insular lobe
The main surface of the cloak lobes consists of grooves and convolutions. The grooves (sulci) are deep folds of the mantle containing stratifiedly located neuron bodies - the cortex (gray matter of the mantle) and cell processes (white matter of the mantle). Between these grooves there are rollers of the cloak, which are usually called convolutions. They contain the same components as the grooves. Each section has its own permanent grooves and convolutions.
The grooves of the telencephalon are divided into three main categories, which reflect their depth, occurrence and stability of outline.
Constant (main) grooves (first order grooves). A person has 10 of them. These are the deepest folds on the surface of the brain, which change least among different people. First order furrows arise in the process early development and are characteristic of every species of animal and human.
Non-permanent grooves (furrows of the second order). These folds, located on the surface of the telencephalon hemispheres, have a characteristic location and direction in which they are oriented. These grooves can individually vary within very wide limits or even be absent. The depth of these grooves is quite large, but significantly less than that of the first-order grooves.
Non-permanent grooves (third order grooves) are called sulci. They rarely reach significant sizes, their outlines are variable, and the topology has ethnic or individual characteristics. As a rule, third-order furrows are not inherited.
The shape of the grooves and convolutions has great individual variability and is a visual criterion (comparable to a fingerprint pattern) that distinguishes one person from another.
The grooves and convolutions are convenient to study on the surface of the telencephalon hemispheres.
The telencephalon is divided by a longitudinal fissure into two hemispheres, connected to each other through a system of commissures. The cerebral hemispheres are the most progressively developing part of the brain in vertebrates. During the evolution of vertebrates, the hemispheres become relatively and absolutely larger. Even in such primitive placental animals as the hedgehog, they dominate the brain. If the total mass of the brain is taken as 1000, then in an elephant the hemispheres account for 630/1000, and the brain stem accounts for 370/1000, in a cat - 614/1000 and 386/1000, respectively, in narrow-nosed monkeys (for example, monkeys) - 709/1000 and 291/1000. Finally, in humans the hemispheres are 780/1000 total mass brain, and for all other parts of the brain, including the cerebellum - only 220/1000.
Each hemisphere is divided into 5 lobes: frontal, temporal, parietal, occipital and insular. In humans, the frontal lobe of the hemisphere weighs 450 g, the parietal lobe - 251 g, the temporal and occipital lobe together - 383 g.
The cerebral hemispheres have a complex topography due to the presence of grooves and convolutions. The surface of the hemispheres is covered with gray matter - the cerebral cortex. The inner parts of the hemispheres consist of white matter, in which the nerve nuclei and lateral ventricles are located.
Cortex of the hemispheres
The cerebral cortex is the most differentiated and complex neural structure. Associated with the bark higher forms reflections of the external world, all types of conscious human activity.
The formation of hemispheric furrows begins in the 5th month of embryonic development. The first to form is the lateral (Sylvian) fissure, then other primary fissures appear: central (Rolandic), parieto-occipital, hippocampal, calcarine. From 7 months, the process of furrow formation proceeds very quickly, secondary furrows appear and at the end of the intrauterine period, the relief of the hemispheres is mainly formed. After birth, small tertiary grooves form, which determine the individual characteristics of the relief of the hemispheres. There are differences in the location of the grooves of the right and left hemispheres, as well as in the size of some gyri. It is believed that the differences serve as an external expression of the functional asymmetry of the cerebral hemispheres.
The surface area of both hemispheres varies in adults from 1469 to 1670 cm2. Of the total surface of the cortex, 2/3 is located deep in the grooves and fissures, and 1/3 is occupied by the convolutions and the visible surface of the hemispheres. In humans, the thickness of the cortex ranges from 1.25 to 4 and even up to 6 mm. In the depths of the grooves, the width of the cortex decreases by 2-2.5 times compared to the apex of the gyrus. If we take into account that the surface of the cortex in one hemisphere is on average 800 cm 2, and its thickness is on average 3 mm, then the volume of the cortex will be 240 cm 3, or 44% of the total volume of the hemisphere. The number of cortical neurons is estimated at 10-18 billion, their total mass is 21.5 g, and their volume is about 20 cm 3 (1:27 relative to the volume of the cortex). If we assume that the processes of one neuron have an average length of 4 cm, then the length of all nerve fibers in the cortex will exceed 400,000 km.
The doctrine of the structure of the cerebral cortex, its architectonics, has several sections. Neuroarchitectonics, or cytoarchitectonics, studies the neuronal composition of the cortex, myeloarchitectonics examines its fibrous structure, glioarchitectonics deals with glial elements, angioarchitectonics deals with the distribution of blood vessels in the cortex.
In phylogenetic terms, the ancient (paleocortex), old (archeocortex) and new (neocortex) bark are distinguished. The ancient and old cortex are located on the medial and basal surface of the hemisphere. They are surrounded by interstitial cortical formations, identified as peripaleocortex and periarchicortex (mesocortex). The ancient bark accounts for only 0.6% of the total crust surface, the old bark - 2.2%, and the intermediate bark - 1.6%. Collectively, this represents 4.4% of the hemisphere's surface area. The remaining 95.6% of the surface is occupied by new crust.
Based on embryonic development, the ancient, old and interstitial cortex belong to the heterogenetic cortex, which does not go through the stage of a six-layer structure and in its final form has a smaller number of layers. The neocortex is defined as the homogenetic cortex. At the 6th month of intrauterine development, its rudiment is divided into 6 layers. In the future, the six-layer bark may remain. In this case, they speak of a homotypic cortex. If after the six-layer stage the number of layers increases or decreases, then such a cortex is called heterotypic.
The surface layer of the homotypic homogenetic cortex is called molecular plate. It consists of a plexus of nerve fibers and contains a few horizontal neurons. The thickness of this layer is 0.15-0.20 mm. The second layer forms outer granular plate 0.10-0.16 mm thick with densely located small granular neurons. It also contains small pyramidal neurons. The third layer is called outer pyramidal plate, its thickness is 0.8-1.0 mm. It is characterized by the presence of medium and large pyramidal neurons with long axons. Deeper lies internal granular plate, which contains small granular and stellate neurons. The width of this layer is 0.12-0.30 mm. The fifth layer is represented internal pyramidal plate thickness 0.4-0.5 mm. Here are the largest pyramidal neurons with axons extending beyond the cortex. The sixth layer is multiform plate, in which neurons of various shapes are located. Its width is 0.6-0.9 mm. The three outer layers of the cortex are usually united under the name of the main outer zone, the three inner ones - under the name of the main inner zone.
The heterotypic cortex differs from the homotypic one in that the internal granular plate is weakly expressed or absent (agranular cortex). The internal pyramidal plate may be absent. In other places, the outer pyramidal, internal granular or internal pyramidal plates are highly developed and sublayers are distinguished in them.
The functional significance of the cortical plates is determined by their cellular composition and interneuronal connections. Fibers from other layers of the cortex and the opposite hemisphere end in the molecular plate. Here are the branches of the apical dendrites of pyramidal neurons, to which impulses coming from other cortical neurons are switched. The external granular and external pyramidal plates contain mainly associative neurons that carry out intracortical connections horizontally and vertically. These two plates represent the youngest phylogenetic structures; their strong development is characteristic of the human cerebral cortex. In ontogenesis, the external granular and external pyramidal plates differentiate and mature later than the others. The internal granular plate is the main receptive layer of the cortex. Most of the specific projection afferent fibers from the thalamus and geniculate nuclei terminate here. The internal pyramidal plate is the origin of the efferent projection pathways. The lamina multiforme contains functionally heterogeneous neurons. It is believed that commissural fibers depart from them, as well as fibers heading to the overlying cortical layers.
Along with the horizontal organization of the cortex in the form of plates, the principle of vertical modular organization of the cortex is currently being considered. The modules are based on such structural components as columns of neurons and bundles of their apical dendrites. It is generally accepted that in the cerebral cortex there are two types of stable genetically determined associations of neurons: micro- and macrocolumns. In the process of life activity, functionally mobile and structurally varying neuron modules can be formed from them.
The microcolumn is considered the main modular subunit in the cortex. It is a vertically oriented strand of cells consisting of approximately 110 neurons and passing through all the plates of the cortex. Cortical columns are modules, information processing units that have their own input and output. The diameter of the columns is about 30 µm. In almost all areas of the cortex, the number of neurons in the columns is relatively constant, and only in the cortical visual centers the number of neurons in the columns is greater. Several hundred microcolumns are combined into a larger structure - a macrocolumn, having a diameter of 500 to 1000 μm. The cortical columns are surrounded by radially arranged nerve fibers and blood vessels. Each such module is considered as a focus of convergence of several thousand local, associative and callosal fibers. There are topographically ordered neural connections between the cortical columns and subcortical formations; certain groups of neurons in the basal ganglia, thalamus, and geniculate bodies correspond to individual columns.
The simplest and most constant associations of neuron elements are bundles of dendrites. Vertical bundles of dendrites appear to play the main constructive role in the consolidation of neurons. The connection of neurons of various microcolumns is carried out mainly by axon terminals of relay efferent fibers, and of macrocolumns by associative and callosal fibers.
Individual dendrites in a bundle can be directly adjacent to each other over a considerable distance, which creates favorable conditions for the implementation of non-synaptic influences of the exchange of ions and metabolites. In associations of neurons formed with the help of bundles of dendrites, structural prerequisites are created for both divergence and convergence of synaptic impulses.
From the point of view of myeloarchitectonics, radial and tangential nerve fibers are distinguished in the cortex. The first enter the cortex from the white matter, or vice versa, exit the cortex into the white matter. The latter are located parallel to the surface of the cortex and form plexuses called stripes at a certain depth. There are strips of the molecular plate, outer and inner granular plates, and internal pyramidal plate. Fibers running in the stripes connect neurons of adjacent cortical columns with each other. The number of stripes in different fields of the cortex is not the same. Depending on it, single-strip, double-strip and multi-strip types of cortex are distinguished. The stripes are especially well defined in the occipital lobe, in the visual fields (striate cortex).
IN nervous system The principle of unity of structure and function is especially clear. At one time, I.P. Pavlov formulated the principle of structure in relation to the nervous system, according to which everything the smallest details brain structures have dynamic (functional) significance. Based on this, it must be recognized that the features of the architectonics of the cerebral cortex, differences in the structure of its areas and fields are associated with their functional functions.
In structural and functional terms, the cerebral cortex can be divided into anterior (frontal lobe) and posterior (occipital, parietal and temporal lobes) sections. The border between them runs along the central groove. The posterior section perceives afferent signals. The cortical fields located here are unequal in functional terms, and they can be divided into primary, secondary and tertiary.
Primary cortical fields are clearly demarcated areas that correspond to the central parts of the analyzers. The bulk of signals from the sensory organs pass into these fields along specific projection afferent pathways. Primary fields are characterized by strong development of the internal granular plate. Primary fields are associated with the relay nuclei of the thalamus and the nuclei of the geniculate bodies. They have a screen structure and, as a rule, a rigid somatotopic projection, in which individual areas of the periphery are projected into the corresponding areas of the cortex. Damage to the primary fields of the cortex is accompanied by a violation of direct perception and fine differentiation of stimuli.
The representation of cutaneous and conscious proprioceptive sensitivity is located in the primary cortical fields (1, 2, 3), occupying the postcentral gyrus. In each hemisphere there is a reverse projection of the surface of the opposite half of the body. In the upper part of the gyrus there is a projection of the lower limb, below is a projection of the abdomen, chest, and even lower is the lower limb. Most bottom part The postcentral gyrus is occupied by zones associated with the innervation of the head and neck, but the projection of parts of the face is not reverse, but direct. Data from studying the columnar organization of the cortex indicate that each body segment (dermatome) is projected onto the cortex in the form of a narrow strip running from front to back through all the cytoarchitectonic fields of the postcentral cortex; in this case, afferent fibers from the skin approach the columns of field 1, field 2 - from the joints, and field 3 - from the muscles.
The primary visual field (17) is located on the medial surface of the hemisphere along the calcarine sulcus. Here the internal granular plate is well developed, which is divided into 3 parts by white stripes. The cortical columns form alternating vertical plates for the right and left eyes. It is believed that neurons in the deep layers of the cortex have the properties of a “motion analyzer,” and neurons in the superficial layers act as a “shape analyzer.”
Primary auditory fields (41, 42) are localized in the transverse temporal gyrus (Heschl) and enter the superior temporal gyrus. These fields represent in order the areas of the cochlea that perceive different sound frequencies. The division into columns is most clearly expressed in the auditory cortex.
The primary olfactory fields are found in the archeocortex, which covers the olfactory triangle, the anterior perforated substance, the septum pellucidum, and the subcallosal field.
The primary taste field is located, according to most researchers, in the lower part of the postcentral gyrus, in the depth of the lateral sulcus, and corresponds to the projection of the tongue.
The cortical end of the vestibular analyzer, according to various authors, has a representation in the middle temporal gyrus (area 21), the superior parietal lobule, and the superior temporal gyrus.
The representation of internal organs in the cortex has not been sufficiently studied; apparently, it is more diffuse in nature. An important role in regulating the functions of the viscera is played by the limbic region of the cortex, which includes the cingulate and parahippocampal gyri, hippocampus, septum pellucidum and subcallosal area. The limbic cortex, together with the subcortical formations (amygdala, nucleus of the leashes, nuclei of the mammillary bodies) makes up limbic system, which represents the substrate of emotions and reactions associated with basic biological drives (hunger, thirst, fear, etc.).
Secondary cortical fields adjacent to the primary fields. They can be considered as peripheral parts of cortical analyzers. These fields are associated with the association nuclei of the thalamus. When secondary fields are damaged, elementary sensations are preserved, but the ability to more complex perceptions is impaired. Secondary fields do not have clear boundaries, and the somatotopic projection is not expressed in them.
The secondary field of general sensitivity is localized in the superior parietal lobule (fields 5, 7). Secondary visual fields (18, 19) occupy the medial surface of the occipital lobe and most of the lateral surface. The secondary auditory field (22) is located in the superior and middle temporal gyri. Secondary olfactory and gustatory fields are localized in the parahippocampal gyrus and uncus (fields 28, 34).
Tertiary cortical fields are distinguished by the finest neural structure and the predominance of associative elements. They occupy the entire inferior parietal lobule and part of the superior parietal lobule, as well as the occipital-temporo-parietal region. These fields are associated with the posterior nuclei of the thalamus. In the tertiary fields, the most complex interactions of analyzers are carried out, underlying the cognitive process (gnosis), and programs of purposeful actions are formed (praxia).
The temporal lobe cortex is involved in storing and reproducing impressions. With electrical stimulation of certain points of the temporal cortex, peculiar reactions are observed in the form of “outbursts of what has been experienced” or a feeling of “already seen.” It is believed that a neural recording of the stream of consciousness is created in the cortex of the temporal lobes; it is stored indefinitely, but cannot be reproduced voluntarily, and “comes to life” only with artificial stimulation and certain painful conditions.
The anterior hemisphere is related to the organization of actions and is also divided into primary, secondary and tertiary cortical fields. The primary motor field (4) is located in the precentral gyrus. There is no internal granular lamina (agranular cortex) and the internal pyramidal lamina with Betz's giant pyramidal neurons is especially strongly developed. The axons of these neurons form a pyramidal tract. Betz cells are directly affected by impulses coming from the cerebellum through the central medial nucleus of the thalamus. In the primary motor field, all the muscles of the body are represented in a reverse projection, as is the skin in the postcentral gyrus. The cortex here is divided into columns that are connected to specific groups of motor neurons in the spinal cord and control the movement of individual muscles or muscle groups.
Secondary motor fields (6, 8) are located anterior to the precentral gyrus. They are characterized by strong development of the outer and inner pyramidal plates, in which large pyramidal neurons predominate. Signals from the cerebellum are transmitted to the secondary fields. Efferent fibers go from here to the nuclei of the striatum. Thus, secondary motor fields are related to the extrapyramidal system; their function is necessary for the performance of complex stereotypic motor acts. Primary and secondary motor fields have rich connections with the posterior hemisphere. Feedback between the movement apparatus and the cortex is carried out through the cerebellum, which perceives proprioceptive stimuli and, after appropriate processing, transmits them to the cerebral cortex.
Tertiary fields occupy most of the frontal lobe, accounting for about 1/4 of the entire surface of the cortex. Here the internal granular plate is well defined, to the neurons of which fibers from the medial nuclei of the thalamus go. The tertiary fields of the frontal cortex are associated with higher forms of purposeful activity and play important role in social behavior. When they are damaged, sensation or movement is not impaired, but the person becomes passive, cannot evaluate current events and his behavior, and loses the ability to foresight.
The most important feature of a person is articulate speech. Academician I.P. Pavlov attributed the speech to the second signaling system, with the help of which there is an indirect reflection of reality. Speech functions are widely represented in the cerebral cortex. Based on data obtained from electrical stimulation and removal of various areas of the cortex in patients, three cortical speech fields were identified. The posterior speech field is located in the occipital-temporo-parietal region, encompassing all three temporal, supramarginal and angular gyri. This field is primarily associated with the perception and understanding of speech and is functionally the leading one. When it is damaged, speech disorder always occurs - aphasia. The anterior speech field lies in the posterior part of the inferior frontal gyrus and corresponds to Broca's motor speech center. The superior, additional, speech field is localized at the upper edge of the hemisphere anterior to the precentral gyrus; when it is damaged, speech disorders are not always observed. Speech fields, like other parts of the cortex, are connected to the nuclei of the thalamus. The posterior field is connected with the posterior nucleus, the superior field with the lateral nucleus, and the anterior field with the medial nuclei. All speech fields are connected by associative paths into a single functional system.
A feature of the speech centers of the cortex is their asymmetry. In most people, they are localized in the left hemisphere, which is dominant in relation to speech. It is generally accepted that this dominance is associated with right-handedness, and that in left-handed people speech is controlled by right hemisphere. Recently, the issue of functional asymmetry of the hemispheres has been interpreted more widely. The left hemisphere is associated with speech and abstract thinking, and the right hemisphere is associated with spatial understanding, imaginative thinking, and musical abilities.
Finite brain develops from the anterior cerebral bladder, consists of highly developed paired parts - the right and left hemispheres and the middle part connecting them.
Rice The furrows and convolutions of the left hemisphere of the cerebrum; superolateral surface
(enlarge picture)
The hemispheres are separated by a longitudinal fissure, in the depth of which lies a plate of white matter, consisting of fibers connecting the two hemispheres - the corpus callosum. Under the corpus callosum there is a vault, which consists of two curved fibrous cords, which are connected to each other in the middle part, and diverge in front and behind, forming the pillars and legs of the vault. Anterior to the columns of the arch is the anterior commissure. Between the anterior part of the corpus callosum and the fornix is a thin vertical plate of brain tissue - a transparent septum.
The hemisphere is formed by gray and white matter. It contains the largest part, covered with grooves and convolutions - a cloak formed by the gray matter lying on the surface - the cortex of the hemispheres; the olfactory brain and accumulations of gray matter inside the hemispheres - the basal ganglia. The last two sections constitute the oldest part of the hemisphere in evolutionary development. The cavities of the telencephalon are the lateral ventricles.
In each hemisphere there are three surfaces: superolateral (upper lateral) convex according to the cranial vault, middle (medial) - flat, facing the same surface of the other hemisphere, and lower - irregular in shape. The surface of the hemisphere has complex drawing, thanks to those going to various directions furrows and ridges between them - convolutions. The size and shape of the grooves and convolutions are subject to significant individual fluctuations. However, there are several permanent grooves that are clearly expressed in everyone and appear earlier than others during the development of the embryo.
They are used to divide the hemispheres into large areas called lobes. Each hemisphere is divided into five lobes: the frontal, parietal, occipital, temporal and hidden lobe, or insula, located deep in the lateral sulcus. The boundary between the frontal and parietal lobes is the central sulcus, and between the parietal and occipital lobes is the parieto-occipital sulcus. The temporal lobe is separated from the rest by the lateral sulcus. On the superolateral surface of the hemisphere in the frontal lobe, there is a precentral sulcus, separating the precentral gyrus, and two frontal sulci: superior and inferior, dividing the rest of the frontal lobe into the superior, middle and inferior frontal gyri.
In the parietal lobe there is a postcentral sulcus, separating the postcentral gyrus, and an intraparietal sulcus, dividing the rest of the parietal lobe into the superior and inferior parietal lobes. In the lower lobule, the supramarginal and angular gyri are distinguished. In the temporal lobe, two parallel grooves - the superior and inferior temporal - divide it into the superior, middle and inferior temporal gyri. In the region of the occipital lobe, transverse occipital sulci and gyri are observed. On the medial surface, the sulcus of the corpus callosum and the cingulate are clearly visible, between which the cingulate gyrus is located.
Above it, surrounding the central sulcus, lies the paracentral lobule. Between the parietal and occipital lobes the parieto-occipital groove passes, and behind it there is the calcarine groove. The area between them is called a wedge, and the one lying in front is called a pre-wedge. At the point of transition to the lower (basal) surface of the hemisphere lies the medial occipitotemporal, or lingual, gyrus. On the lower surface, separating the hemisphere from the brain stem, there is a deep groove of the hippocampus (seahorse groove), lateral to which is the parahippocampal gyrus. Laterally, it is separated by a collateral groove from the lateral occipitotemporal gyrus. The insula, located deep in the lateral (side) sulcus, is also covered with grooves and convolutions.
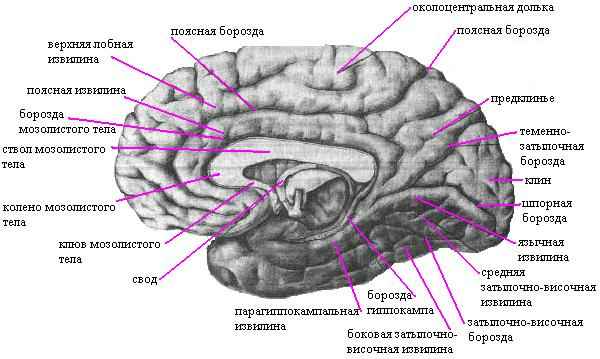 Cerebral cortex It is a layer of gray matter up to 4 mm thick. It is formed by layers of nerve cells and fibers arranged in a certain order.
Cerebral cortex It is a layer of gray matter up to 4 mm thick. It is formed by layers of nerve cells and fibers arranged in a certain order.
Rice. The furrows and convolutions of the right hemisphere of the cerebrum; medial and inferior surfaces.
(enlarge picture)
The most typically structured areas of the phylogenetically newer cortex consist of six layers of cells; the old and ancient cortex has fewer layers and is simpler in structure. Different areas of the cortex have different cellular and fibrous structures. In this regard, there is a doctrine about the cellular structure of the cortex (cytoarchitectonics) and the fibrous structure (myeloarchitectonics) of the cerebral hemisphere cortex.
Olfactory brain in humans it is represented by rudimentary formations, well expressed in animals, and constitutes the oldest parts of the cerebral cortex.
Basal ganglia are accumulations of gray matter within the hemispheres. These include the striatum, consisting of the caudate and lenticular nuclei, interconnected. The lenticular nucleus is divided into two parts: the shell, located on the outside, and the globus pallidus, which lies on the inside. They are subcortical motor centers.
Outside the lenticular nucleus there is a thin plate of gray matter - a fence, in anterior section The temporal lobe contains the amygdala. Between the basal ganglia and the optic thalamus there are layers of white matter, the inner, outer and outermost capsules. Conducting pathways pass through the internal capsule.
Lateral ventricles(right and left) are cavities of the telencephalon, lie below the level of the corpus callosum in both hemispheres and communicate through the interventricular foramina with the third ventricle. They have an irregular shape and consist of anterior, posterior and lower horns and a central part connecting them. The anterior horn lies in the frontal lobe; it continues posteriorly into the central part, which corresponds to the parietal lobe. At the back, the central part passes into the posterior and inferior horns, located in the occipital and temporal lobes. In the lower horn there is a cushion - the hippocampus (seahorse). From the medial side, the choroid plexus invaginates into the central part of the lateral ventricles, continuing into the inferior horn. The walls of the lateral ventricles are formed by the white matter of the hemispheres and the caudate nuclei. The thalamus is adjacent to the central part below.
The white matter of the hemispheres occupies the space between the cortex and the basal ganglia. It consists of large quantity nerve fibers going to different directions. There are three systems of fibers of the hemispheres: associative (combinative), connecting parts of the same hemisphere; commissural (commissural) connecting parts of the right and left hemispheres, which in the hemispheres include the corpus callosum, anterior commissure and commissure of the fornix, and projection fibers, or pathways connecting the hemispheres with the underlying parts of the brain and spinal cord.



