Úloha mozkových hemisfér. Stavba a význam mozkových hemisfér
1. Zapamatujte si ze svého kurzu zoologie, jaké části mozku mají všichni obratlovci. Deriváty kterého oddělení jsou mozkové hemisféry? Ve které skupině zvířat se objevují poprvé?
Všichni obratlovci se vyznačují přítomností 5 částí mozku: medulla, zadní mozek, střední mozek, diencephalon, přední mozek.
Mozkové hemisféry jsou deriváty předního mozku a objevují se nejprve u obojživelníků, ale u nich jsou špatně vyvinuté a mozková kůra prakticky chybí.
2. Popište strukturu mozkové hemisféry osoba. Nakreslete schematický příčný řez mozkovými hemisférami s vyznačením šedé hmoty kůry, šedé hmoty jader, bílé hmoty a komor.
U člověka je přední mozek reprezentován dvěma hemisférami a corpus callosum, které spojuje hemisféry. Mozkové hemisféry: pravá a levá - pokrývají střední mozek a diencephalon a tvoří až 80 % hmoty dospělého mozku. Na povrchu každé polokoule je mnoho rýh a záhybů. Existují 4 hlavní sulci (centrální, laterální a parietookcipitální), které rozdělují každou hemisféru na laloky. Povrchovou vrstvu tvoří šedá hmota (kortex), pod ní je bílá hmota, tvořená axony nervových buněk, jejichž těla leží v kůře nebo které přenášejí informace do kortikálních buněk. V tl bílá hmota lhát velké shlukyšedá hmota (subkortikální jádra) a dutina (laterální komory). Podél mozkových komor a centrální kanál Míchou cirkuluje mozkomíšní mok, který zajišťuje výživu podkorových struktur.
3. Co je mozková kůra velký mozek? Kde to je?
Povrchová vrstva šedé hmoty mozkových hemisfér se nazývá kůra. Kůra se skládá z několika vrstev neuronových těl, které se liší strukturou a funkcí. Předpokládá se, že se skládá z asi 12-18 miliard buněk, tloušťka je 1,5 - 4,5 mm a plocha je 1,7 - 2,5 tisíc cm2.
4. Vysvětlete význam rýh a záhybů na povrchu mozkových hemisfér.
Brázdy a konvoluce výrazně zvětšují povrch mozkové kůry, podle některých zdrojů až 10-12krát.
5. Jakou funkci plní bílá hmota mozkových hemisfér?
Bílá hmota tvoří cesty, které spojují oblasti kůry a kůry s dalšími sekcemi nervový systém.
6. Jaké laloky se rozlišují v mozkových hemisférách?
Hemisféry se dělí na čelní, 2 parietální, 2 temporální a týlní laloky
7. Rozlišujte pojmy „laloky mozkových hemisfér“ a „zóny mozkových hemisfér“. Uveďte příklady, kdy se shodují nebo neshodují.
Laloky mozkových hemisfér jsou rozdělením povrchu kůry podle anatomického principu: v každé hemisféře jsou čelní, okcipitální, parietální a temporální laloky.
Kortikální zóny jsou úsekem mozkové kůry, vyznačující se jednotností struktury a funkcí.
V našem mozku se laloky a zóny neshodují, jeden lalok se skládá z několika zón. Týlní lalok se skládá z oblastí vidění a vizuálního rozpoznávání. Oblasti čichu, sluchu a chuti se nacházejí ve spánkovém laloku a přiléhají k němu.
8. Při vyšetření nevidomého pacienta bylo zjištěno, že jeho oči a zrakové nervy nejsou poškozeny. Proč pořád nevidí?
Neschopnost vidět může být způsobena nejen poškozením očí nebo zrakových nervů, ale také poškozením zrakových oblastí týlních laloků mozkových hemisfér.
9. Pomocí dalších zdrojů informací zjistěte, zda se liší funkce levé a pravé hemisféry velkého mozku.
Mezi pravou a levou hemisférou mozku existuje „funkční asymetrie“, to znamená, že jejich funkce jsou odlišné. To se prokázalo při pokusech o přerušení komunikačních cest mezi hemisférami (následně se takové operace začaly provádět z lékařských důvodů u některých nemocí, např. u Parkinsonovy choroby). U praváků je vedoucí hemisféra levá, u leváků je vedoucí hemisféra pravá. Pravá hemisféra je zodpovědný za kreativní myšlení, tvoří základ kreativity, dělá nestandardní rozhodnutí; Vizuální oblast pravé hemisféry je zodpovědná za rozpoznávání tváří. Levá hemisféra poskytuje logické uvažování a abstraktní myšlení, obsahuje centra ústní a písemné řeči a utváření rozhodnutí; Vizuální oblast je zodpovědná za rozpoznávání písmen a číslic. Proto, jako vtip, pravákům se říká matematici a levákům se říká umělci.
10. Existují každodenní pojmy „mužská logika“ a „ženská logika“. Existuje nějaký základ pro takové rozdíly?
Ve vnějších a vnitřní struktura Mezi mozky mužů a žen není rozdíl, ale i přes to, že struktura našeho mozku je téměř stejná, jsme individuální. Jsou uvedeny hlavní rozdíly ve vnímání světa podle „mužského“ nebo „ženského“ typu raného dětství v závislosti na naší výchově a vlivem gonadálních hormonů během dospívání, ale všechny jsou pouze psychického rázu.
Mozkové hemisféry jsou nejvyšším oddělením centrálního nervového systému. Toto je největší část mozku. Jsou to párové útvary spojené corpus callosum, což je vlákno nervových vláken. U dospělého člověka tvoří mozkové hemisféry až 80 % mozkové hmoty. Nahoře jsou pokryty šedou hmotou - mozková kůra. V kůře je 12 -18 miliard nervových buněk. Povrch mozkové kůry u dospělého člověka dosahuje 2200-2600 cm2. Podívejme se v tomto článku na strukturu a význam mozkových hemisfér.
To je zajímavé. Platí, že čím větší mozek, tím vyšší inteligence jeho majitele? Není pochyb, že čím více nervových buněk („šedé hmoty“) je v mozku, tím složitější může být spojení mezi nimi a tím vyšší inteligence člověk může mít. Ale zároveň samotná velikost nestačí. Důležitější je organizace mozku. Například, vynikající spisovatel Ivan Turgenev měl mozek o hmotnosti 1700 g a neméně vynikající Anatole France - pouze 1100 g. Průměrná hmotnost lidského mozku je přitom 1400-1500 g.
Téměř všechny dovednosti, které člověk získá během života, tak či onak souvisí s funkcemi mozkové kůry. Kůra je materiální základ psychika. Poskytuje řeč, duševní činnost a paměť.
Četné rýhy (prohlubně) rozdělují polokoule na konvexní konvoluce (záhyby) a laloky. Složená struktura výrazně zvětšuje povrch a objem kůry.
Tři hlavní brázdy - centrální, postranní A parietookcipitální- každá hemisféra mozku je rozdělena na čtyři laloky: frontální, parietální, okcipitální a temporální (obr.). Laloky jsou zase rozděleny drážkami do mnoha záhybů.
Rýže. Mozková kůra
Různé části mozkové kůry plní různé funkce, proto jsou rozděleny do zón. Existují zóny senzorické (senzitivní), asociativní a motorické (motorické).
Smyslové oblasti jsou nejvyšší centra různé typy citlivost. Při jejich podráždění vznikají čití, při poškození dochází k narušení smyslových funkcí (slepota, hluchota atd.).
V okcipitální oblasti kůry se nachází zraková, v časové - čichová, chuťová a sluchová senzorická zóna (obr.).
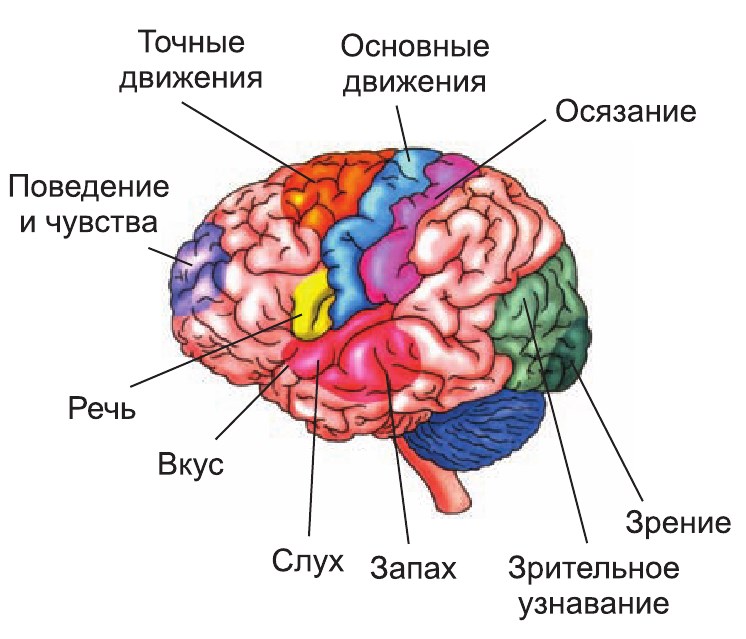
Rýže. Lokalizace funkcí v mozkové kůře
Zóny kožních a svalových smyslů jsou umístěny za centrálním sulkem a motorická zóna je před ním. Nejvíc velké velikosti mají smyslové oblasti rukou a obličeje. A to je vzhledem k důležitosti těchto orgánů v životě člověka pochopitelné. Nejmenší velikosti na senzorické oblasti trupu, stehna a bérce.
Když impulsy vstupují do smyslových zón, dochází také k excitaci asociativní. Kromě toho může stimulace přicházet do stejné asociativní zóny z různých smyslových orgánů. Například ve zrakové asociativní zóně se excitace objevuje nejen jako odpověď na vizuální, ale také na sluchovou stimulaci. Při narušení funkcí asociativních zón ztrácí člověk schopnost správně vyhodnocovat jevy a děje.
Zvláště důležitá role V komplexních formách chování hrají roli frontální asociativní zóny mozkové kůry. Poskytují zpracování smyslových informací a tvoří cíl a program činnosti. Program se skládá z týmů, které jsou vysílány výkonným orgánům. Z nich se vrací informace do frontálních asociačních zón, kde se zjišťuje, zda bylo cíle dosaženo či nikoliv. V druhém případě je příkaz upraven. Vývoj těchto laloků kůry je do značné míry spojen vysoká úroveň duševní schopnosti lidí ve srovnání se zvířaty.
Motorové zóny se nazývají úseky mozkové kůry, které řídí dobrovolné pohyby. Motorická funkce různých částí těla je zastoupena v předním centrálním gyru. Největší prostor zabírají ho motorické zóny rukou, prstů a obličejových svalů a nejméně svaly trupu.
Funkce hemisfér
Správně a levá hemisféra provádět u lidí různé funkce. V vlevo, odjet Hemisféry obsahují centra ústní a písemné řeči. Zde se provádějí procesy analýzy a syntézy informací, zobecnění a rozhodnutí. Verbálně-logické myšlení poskytované levou hemisférou umožňuje člověku poznat podstatu předmětu a jít dál individuální svět. Na jejím základě se tvoří lidské poznání. Předává se z generace na generaci zaznamenáváním verbálních nebo symbolických signálů.
Že jo hemisféra provádí imaginativní myšlení. Pracuje s obrazy objektů ve vnějším světě a dokáže z nich vytvořit nevídané, fantastické kombinace. A to je základ kreativity a nevšedních rozhodnutí. Význam pravé hemisféry pro hudební a umělecká tvořivost. Je známo, že nejvíce vynikající umělci, básníci, hudebníci jsou lidé s převahou pravohemisférového myšlení.
I přes funkční asymetrie, mozek funguje jako celek. Shrnováním informací zajišťuje adekvátní chování, myšlení, vědomí, paměť, práci a tvůrčí činnost osoba.
14.1. OBECNÁ USTANOVENÍ
Telencephalon neboli velký mozek, umístěný v supratentoriálním prostoru lebeční dutiny a se skládá ze dvou velkých
hemisféry (gemispherium cerebralis),odděleny od sebe hlubokou podélnou štěrbinou (fissura longitudinis cerebri), do kterého je ponořen falx cerebri (falx cerebri), představující zdvojení dura mater. Mozkové hemisféry tvoří 78 % jeho hmoty. Každá z mozkových hemisfér má laloky: frontální, parietální, temporální, týlní a limbický. Pokrývají struktury diencefala a mozkového kmene a mozečku umístěné pod cerebelárním tentorium (subtentoriální).
Každá z mozkových hemisfér má tři povrchy: superolaterální nebo konvexitní (obr. 14.1a), - konvexní, směřující ke kostem lebeční klenby; vnitřní (obr. 14.1b), přiléhající k velkému falciformnímu výběžku, a dolní neboli bazální (obr. 14.1c), opakující reliéf spodiny lební (její přední a střední jamky) a tentorium mozečku. V každé hemisféře jsou tři okraje: horní, dolní vnitřní a spodní vnější a tři póly: přední (frontální), zadní (týlní) a laterální (temporální).
Dutina každého Cerebrální hemisféračiní postranní mozková komora, v tomto případě je levá boční komora rozpoznána jako první, pravá - jako druhá. Postranní komora má centrální část umístěnou hluboko v parietálním laloku (lobus parietalis) a tři rohy vybíhající z něj: přední roh proniká do čelního laloku (lobus frontalis), nižší - k časovému (lobus temporalis), zadní - do okcipitálního (lobus occipitalis). Každá z postranních komor komunikuje s třetí mozkovou komorou přes interventrikulární Monroeova díra.
Centrální oddělení mediální povrch Obě hemisféry jsou spojeny mozkovými komisurami, z nichž nejmohutnější je corpus callosum, a strukturami diencephalon.
Telencephalon, stejně jako ostatní části mozku, se skládá z šedé a bílé hmoty. Šedá hmota se nachází hluboko v každé hemisféře, tvoří tam podkorové uzliny a podél obvodu volných ploch hemisféry, kde tvoří mozkovou kůru.
Hlavní problémy související se strukturou, funkcemi podkorových uzlin a variantami klinického obrazu při jejich postižení jsou diskutovány v kapitolách 5, 6. Mozková kůra má plochu cca
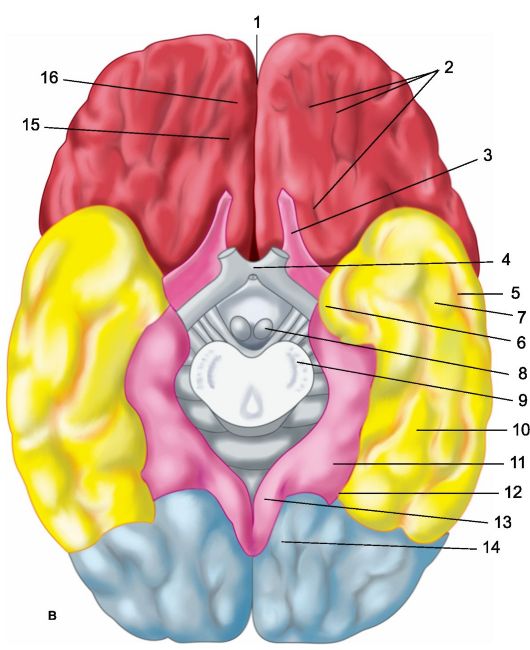
Rýže. 14.1.Hemisféry velkého mozku.
A - superolaterální povrch levá hemisféra: 1 - centrální sulcus; 2 - orbitální část gyrus frontalis inferior; I - čelní lalok; 3 - precentrální gyrus; 4 - precentrální sulcus; 5 - horní frontální gyrus; 6 - střední frontální gyrus; 7 - tegmentální část dolního frontálního gyru; 8 - dolní čelní gyrus; 9 - boční drážka; II - parietální lalok: 10 - postcentrální gyrus; 11 - postcentrální sulcus; 12 - intraparietální sulcus; 13 - supramarginální gyrus; 14 - úhlový gyrus; III - temporální lalok: 15 - gyrus temporalis superior; 16 - sulcus temporalis superior; 17 - střední temporální gyrus; 18 - střední spánková drážka; 19 - dolní temporální gyrus; IV - týlní lalok: b - mediální plocha pravé hemisféry: 1 - paracentrální lalůček, 2 - precuneus; 3 - parieto-okcipitální rýha; 4 - klín, 5 - lingvální gyrus; 6 - laterální okcipitotemporální gyrus; 7 - parahipokampální gyrus; 8 - háček; 9 - klenba; 10 - corpus callosum; 11 - horní frontální gyrus; 12 - cingulární gyrus; c - spodní plocha mozkových hemisfér: 1 - podélná interhemisferická štěrbina; 2 - orbitální drážky; 3 - čichový nerv; 4 - vizuální chiasma; 5 - střední temporální sulcus; 6 - háček; 7 - dolní temporální gyrus; 8 - mastoidní tělo; 9 - báze mozkového peduncle; 10 - laterální okcipitotemporální gyrus; 11 - parahippokampální gyrus; 12 - kolaterální drážka; 13 - cingulární gyrus; 14 - lingvální gyrus; 15 - čichová rýha; 16 - rovný gyrus.
3násobek povrchu hemisfér viditelného při zevním vyšetření. To je způsobeno skutečností, že povrch mozkových hemisfér je složený a má četné prohlubně - brázdy (sulci cerebri) a nachází se mezi nimi konvoluce (gyri cerebri). Mozková kůra pokrývá celý povrch záhybů a rýh (odtud její další název pallium – plášť), někdy proniká do velké hloubky až do hmoty mozku.
Závažnost a umístění rýh a konvolucí mozkových hemisfér je do určité míry variabilní, ale ty hlavní se tvoří během procesu ontogeneze a jsou konstantní, charakteristické pro každý normálně vyvinutý mozek.
14.2. HLAVNÍ DRÁŽKY A GYRILY HEMISfér MOZKU
Superolaterální (konvexální) povrch hemisfér (obr. 14.1a). Největší a nejhlubší - postranní brázda (sulcus lateralis),nebo sylvian brázda, - odděluje čelní a přední část temenního laloku od dolního spánkového laloku. Čelní a parietální lalok jsou odděleny centrální neboli Rolandický sulcus(sulcus centralis), která protíná horní okraj hemisféry a směřuje podél jejího konvexního povrchu dolů a dopředu, mírně před dosažením laterální sulcus. Parietální lalok je oddělen od týlního laloku umístěného za ním parietookcipitálními a příčnými okcipitálními trhlinami probíhajícími podél mediálního povrchu hemisféry.
Ve frontálním laloku, před centrálním gyrem a rovnoběžně s ním, precentrální (gyrus precentralis), nebo přední centrální, gyrus, který je zepředu ohraničen precentrálním sulkusem (sulcus precentralis). Frontální sulci superior a inferior se rozprostírají vpředu od precentrálního sulku a rozdělují konvexální povrch předních částí frontálního laloku na tři frontální gyry – horní, střední a dolní (gyri frontales superior, media et inferior).
Přední část konvexitní plochy parietálního laloku je tvořena postcentrálním sulkem umístěným za centrálním sulkem. (gyrus postcentralis), nebo zadní centrální, gyrus. Vzadu je ohraničen postcentrálním sulkem, ze kterého směrem dozadu vybíhá intraparietální sulcus. (sulcus intraparietalis), oddělující horní a dolní parietální lalok (lobuli parietales superior et inferior). V dolním parietálním laloku se zase rozlišuje gyrus supramarginální (gyrus supramarginalis), obklopující zadní část laterální (Sylviánské) štěrbiny a úhlový gyrus (girus angularis), hraničící se zadní částí gyru temporalis superior.
Na konvexálním povrchu okcipitálního laloku mozku jsou rýhy mělké a mohou se výrazně lišit, v důsledku čehož je také proměnná povaha mezi nimi umístěných záhybů.
Konvexální plocha spánkového laloku je rozdělena sulcus temporalis superior a inferior, které mají směr téměř paralelní s laterální (Sylviovou) štěrbinou a rozdělují konvexální plochu spánkového laloku na horní, střední a dolní temporální gyri. (gyri temporales superior, media et inferior). Gyrus temporalis superior tvoří spodní ret laterální (Sylviánské) štěrbiny. Na jeho povrchu obklad
straně laterálního sulku je několik příčných malých rýh, které na něm zvýrazňují malé příčné záhyby (Heschlovy konvoluce), které lze vidět pouze roztažením okrajů boční drážky.
Přední část laterální (Sylvovské) štěrbiny je prohlubeň se širokým dnem, tvořící t. zv. ostrov (ostrov), nebo insula (lubus insularis). Horní okraj laterálního sulku pokrývajícího tento ostrůvek se nazývá pneumatika (operculum).
Vnitřní (mediální) povrch hemisféry (obr. 14.1b). Centrální část vnitřního povrchu hemisféry je úzce spojena se strukturami diencephalon, od kterých je oddělena těmi, které se vztahují k velkému mozku. klenba (fornix) A corpus callosum (corpus callosum). Ten je zvenčí ohraničen žlábkem corpus callosum (sulcus corporis callosi), počínaje přední částí - zobákem (řečniště) a končící na jejím zesíleném zadním konci (splenium). Zde rýha corpus callosum přechází do hluboké hipokampální rýhy (sulcus hippocampi), která proniká hluboko do hmoty hemisféry, vtlačuje ji do dutiny dolního rohu postranní komory, čímž vzniká tzv. tzv. amonný roh.
Mírně ustupující od sulcus corpus callosum a hippocampal sulcus se nacházejí kalosální-marginální, subparietální a nazální sulci, které jsou vzájemnými pokračováními. Tyto drážky vymezují vnější obloukovitou část mediálního povrchu mozkové hemisféry, tzv. limbický lalok(lobus limbicus). V limbickém laloku jsou dva gyri. Horní část limbického laloku je horní limbický (superior marginální), neboli obklopující, gyrus (girus cinguli), spodní část tvoří dolní limbický gyrus neboli gyrus mořského koníka (girus hippocampi), nebo parahippokampální gyrus (girus parahyppocampalis), před kterým je háček (strýc).
Kolem limbického laloku mozku jsou útvary vnitřního povrchu čelního, parietálního, okcipitálního a temporálního laloku. Většinu vnitřního povrchu frontálního laloku zabírá mediální strana horního frontálního gyru. Nachází se na hranici mezi frontálním a parietálním lalokem mozkové hemisféry paracentrální lalůček (lobulis paracentralis), který je pokračováním předního a zadního centrálního gyru na mediální ploše hemisféry. Na hranici mezi parietálním a okcipitálním lalokem je jasně viditelný parieto-okcipitální sulcus (sulcus parietooccipitalis). Z jeho spodní části se táhne dozadu kalkarinová drážka (sulcus calcarinus). Mezi těmito hlubokými drážkami je gyrus trojúhelníkového tvaru známý jako klín. (cuneus). Před klínem je čtyřúhelníkový gyrus, související s parietální lalok mozek - precuneus.
Spodní povrch polokoule (obr. 14.1c). Spodní povrch mozkové hemisféry tvoří útvary frontální, temporální a týlní lalok y Část čelního laloku přiléhající ke střední čáře je gyrus rectus (girus rectus). Zvenčí je ohraničena čichovým žlábkem (sulcus olphactorius), ke kterému dole přiléhají útvary čichového analyzátoru: čichový bulbus a čichový trakt. Bočně k ní, až k laterální (Sylviánské) štěrbině, vybíhající na spodní plochu čelního laloku, jsou malé orbitální gyry (gyri orbitalis). Laterální části spodního povrchu hemisféry za laterálním sulkem jsou obsazeny gyrus temporalis inferior. Mediálně k němu je laterální temporo-okcipitální gyrus (gyrus occipitotemporalis lateralis), nebo vřetenovou drážkou. Před-
jeho oddělení s uvnitř hraničí s gyrus hippocampu a ty zadní s lingválem (gyrus lingualis) nebo mediální temporo-okcipitální gyrus (gyrus occipitotemporalis medialis). Ten svým zadním koncem přiléhá ke kalkarinové rýze. Přední úseky vřetenových a lingválních gyri patří do spánkového laloku a zadní úseky do týlního laloku mozku.
14.3. BÍLÁ HMOTA VELKÝCH POLOKUL
Bílá hmota mozkových hemisfér se skládá z nervových vláken, hlavně myelinu, která tvoří dráhy zajišťující spojení mezi kortikálními neurony a shluky neuronů, které tvoří thalamus, subkortikální ganglia a jádra. V jeho hloubce se nachází hlavní část bílé hmoty mozkových hemisfér semioval centrum nebo corona radiata (corona radiata), skládající se převážně z aferentního a eferentního projekce dráhy spojující mozkovou kůru se subkortikálními ganglii, jádry a retikulární substancí diencefala a mozkového kmene, se segmenty míchy. Jsou umístěny zvláště kompaktně mezi thalamem a subkortikálními uzlinami, kde tvoří vnitřní pouzdro popsané v kapitole 3.
Nervová vlákna, která spojují části kůry jedné hemisféry, se nazývají asociativní. Čím kratší jsou tato vlákna a vazby, které tvoří, tím povrchněji jsou umístěny; delší asociativní spojky, umístěné hlouběji, spojují relativně vzdálené oblasti mozkové kůry (obr. 14.2 a 14.3).
Vlákna, která spojují mozkové hemisféry a mají tedy společný příčný směr, se nazývají komisurální, nebo lepidlo. Komisurální vlákna spojují identické oblasti mozkových hemisfér a vytvářejí tak možnost kombinace jejich funkcí. Tvoří se tři komisury velký mozek: nejhmotnější z nich je corpus callosum (corpus callosum), navíc tvoří komisurální vlákna přední komisura, nachází se pod zobákem corpus callosum (rostrum corporis collosum) a spojující obě čichové oblasti, jakož i komisura fornixu (commissura fornicis), nebo hipokampální komisura, tvořená vlákny spojujícími struktury amonných rohů obou hemisfér.
V přední sekce Corpus callosum obsahuje vlákna spojující čelní laloky, dále vlákna spojující temenní a temporální lalok a zadní část corpus callosum spojuje týlní laloky mozku. Přední komisura a fornixová komisura spojují především oblasti starověkého a starého kortexu obou hemisfér, přední komisura navíc poskytuje spojení mezi jejich středním a dolním temporálním gyri.
14.4. OLfactory SYSTEM
V procesu fylogeneze je vývoj velkého mozku spojen s tvorbou čichového systému, jehož funkce přispívají k zachování životaschopnosti zvířat a mají mnoho Důležité pro lidský život.
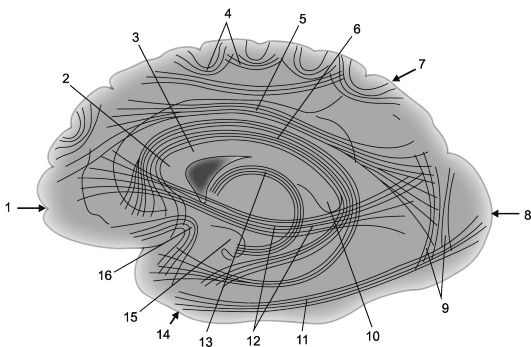
Rýže. 14.2.Asociační kortikálně-kortikální spojení v mozkových hemisférách [podle V.P. Vorobjov].
1 - čelní lalok; 2 - genu corpus callosum; 3 - corpus callosum; 4 - oblouková vlákna; 5 - horní podélný nosník; 6 - cingulární gyrus; 7 - temenní lalok, 8 - okcipitální lalok; 9 - svislé Wernicke nosníky; 10 - splenium corpus callosum;
11 - spodní podélný nosník; 12 - subcallosální svazek (fronto-okcipitální dolní svazek); 13 - klenba; 14 - temporální lalok; 15 - hák gyru hippocampu; 16 - chomáče háčků (fasciculus uncinatus).
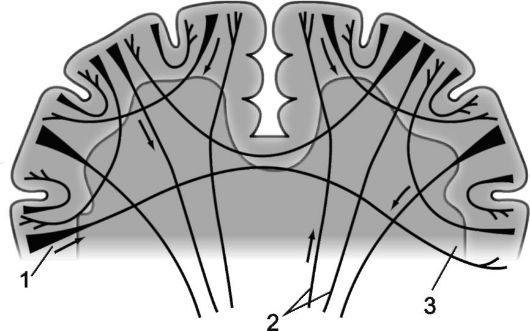
Rýže. 14.3.Myeloarchitektura mozkových hemisfér.
1 - projekční vlákna; 2 - komisurální vlákna; 3 - asociativní vlákna.
14.4.1. Stavba čichového ústrojí
Těla prvních neuronů čichového systému se nacházejí ve sliznici hlavně nos horní část nosní přepážky a horní nosní meatus. Čichové buňky jsou bipolární. Jejich dendrity zasahují až k povrchu sliznice a končí zde specifickými receptory a shluk axonů v tzv čichová vlákna (fili olfactorii), jejichž počet na každé straně je asi dvacet. Takový svazek čichových vláken a tvoří první kraniální neboli čichový nerv(obr. 14.4). Tato vlákna přecházejí do přední (čichové, čichové) lebeční jamky přes etmoidní kost a končí v buňky zde umístěné čichové žárovky. Čichové bulby a proximálně umístěné čichové dráhy jsou ve skutečnosti důsledkem výběžků hmoty velkého mozku, které se tvoří během ontogeneze a představují struktury s ní související.
Čichové bulby obsahují buňky, které jsou těly druhých neuronů čichová dráha, jejíž axony se tvoří čichové dráhy (tracti olfactorii), umístěné pod čichovými rýhami, laterálně k přímým konvolucím umístěným na bazální ploše čelní laloky. Čichové dráhy směřují dozadu do subkortikálních čichových center. Přiblížením k přední perforované ploténce se vlákna čichového traktu dělí na mediální a laterální snopce, tvořící na každé straně čichový trojúhelník. V budoucnu jsou tato vlákna vhodná k tělům třetích neuronů čichového analyzátoru, které se nacházejí
Rýže. 14.4.Čichový analyzátor.
1 - čichové buňky; 2 - čichová vlákna (dohromady tvoří čichové nervy); 3 - čichové žárovky; 4 - čichové dráhy; 5 - čichové trojúhelníky; 6 - parahipokampální gyrus; 7 - projekční zóna čichového analyzátoru (zjednodušené schéma).
v periamygdale a subcallosálních oblastech, v jádrech septum pellucidum, umístěných anteriorně od přední komisury. Přední komisura spojuje obě čichové oblasti a zajišťuje také jejich spojení s limbickým systémem mozku. Některé z axonů třetích neuronů čichového analyzátoru, procházejících přední komisurou mozku, se kříží.
Axony třetích neuronů čichový analyzátor umístěný v podkorových čichových centrech, míří do fylogeneticky stará kůra mediobazální plocha spánkového laloku (k piriformis a parahippokampálním gyri a k uncus), kde se nachází projekční čichová zóna, nebo kortikální konec čichového analyzátoru (pole 28, podle Brodmanna).
Čichový systém je tak jediným smyslovým systémem, ve kterém specifické impulsy obcházejí thalamus na cestě z receptorů do kůry. Ve stejnou dobu čichový systém má zvláště vyjádřené souvislosti s limbickými strukturami mozku a informace jeho prostřednictvím mají významný vliv na stav emoční sféry a funkce autonomního nervového systému. Vůně mohou být příjemné nebo nepříjemné, ovlivňují chuť k jídlu, náladu a mohou způsobit různé autonomní reakce, zejména nevolnost a zvracení.
14.4.2. Studium čichu a významu jeho poruch pro topickou diagnostiku
Při vyšetření čichového stavu je nutné zjistit, zda pacient cítí pachy, zda jsou tyto vjemy na obou stranách stejné, zda pacient rozlišuje povahu vnímaných pachů, zda nemá čichové halucinace - záchvatovité pocity a vůně, která v něm chybí životní prostředí.
Ke studiu čichu využívají pachové látky, jejichž pach není štiplavý (štiplavé pachy mohou způsobit podráždění receptorů trojklaného nervu umístěných v nosní sliznici) a jsou pacientovi známé (v v opačném případě je obtížné rozpoznat perverzi čichu). Čich se kontroluje na každé straně zvlášť, přičemž druhá nosní dírka musí být uzavřena. Můžete použít speciálně připravené sady slabých roztoků pachových látek (máta, dehet, kafr atd.), v praktické práci lze použít i improvizované prostředky (žitný chléb, mýdlo, banán atd.).
Snížený čich - hyposmie, nedostatek čichu - anosmie, zvýšený čich - hyperosmie, perverze pachů - dysosmie, pocit vůně při absenci podnětu - parosmie, subjektivní pocit nepříjemného zápachu, který skutečně existuje a je způsoben organickou patologií v nosohltanu - Cacosmia, neexistující pachy, které pacient paroxysmálně pociťuje - čichové halucinace - jsou častěji čichovou aurou epilepsie temporálního laloku, která může být způsobena různými příčinami, zejména nádorem spánkového laloku.
Hyposmie nebo anosmie na obou stranách je obvykle důsledkem poškození nosní sliznice akutními katarálními stavy, chřipkou, alergickou rýmou, atrofií sliznice
nosu v důsledku chronické rýmy a dlouhodobého užívání vazokonstrikčních nosních kapek. Chronická rýma s atrofií nosní sliznice (atrofická rýma), Sjögrenova choroba odsoudí člověka k přetrvávající anosmii. Bilaterální hyposmie může být způsobena hypotyreózou, diabetes mellitus, hypogonadismem, selháním ledvin, dlouhodobým kontaktem s těžkými kovy, formaldehydem atd.
Ve stejnou dobu jednostranná hyposmie nebo anosmie je často důsledkem intrakraniálního tumoru, nejčastěji meningeomu přední lebeční (čichové) jamky, který tvoří až 10 % intrakraniálních meningeomů, stejně jako některé gliové nádory čelního laloku. Poruchy čichu vznikají v důsledku stlačení čichového traktu na straně patologického ložiska a mohou být po určitou dobu jediným ložiskovým příznakem onemocnění. Vizualizace nádorů může být zajištěna CT nebo MRI skenováním. Se zvětšováním meningeomu čichové jamky se obvykle rozvinou duševní poruchy charakteristické pro frontální syndrom (viz kap. 15).
Jednostranné poškození částí čichového analyzátoru umístěných nad jeho subkortikálními centry v důsledku neúplného křížení drah na úrovni přední mozkové komisury obvykle nevede k výraznému snížení čichu. Podráždění patologickým procesem kůry mediobazálních částí temporálního laloku, především gyrus parahipocampal a jeho uncus, může způsobit záchvatovitý výskyt čichové halucinace. Pacient náhle začne bezdůvodně cítit zápach, často nepříjemného charakteru (zápach spáleniny, shnilého, shnilého, připáleného atd.). Čichové halucinace v přítomnosti epileptogenního ložiska v mediobazálních oblastech spánkového laloku mozku může být projevem aury epileptického záchvatu. Poškození proximální části, zejména kortikálního konce čichového analyzátoru, může způsobit středně těžkou oboustrannou (spíše na opačné straně) hyposmii a zhoršenou schopnost identifikovat a rozlišovat pachy (čichová agnozie). Posledně jmenovaná forma poruchy čichu, která se projevuje ve stáří, je s největší pravděpodobností spojena s dysfunkcí kůry v důsledku atrofických procesů v její projekční čichové zóně.
14.5. LIMBIC-RETIKULÁRNÍ KOMPLEX
V roce 1878 P. Broca(Broca P., 1824-1880) nazývaný „velký okrajový neboli limbický lalok“ (z latinského limbus – hrana) spojoval hippocampus a gyrus cingulate, navzájem spojeny isthmem gyrus cingulate, umístěným nad splenium corpus callosum.
V roce 1937 D. Papets(Papez J.) na základě experimentálních dat vznesl odůvodněnou námitku proti dříve existujícímu konceptu zapojení mediobazálních struktur mozkových hemisfér především na zajišťování čichu. On navrhl, že hlavní část mediobazálních částí mozkové hemisféry, tehdy nazývané čichový mozek (rhinencephalon), ke kterému limbický lalok patří, představuje morfologický základ nervového mechanismu afektivního chování, a sjednotil je pod názvem"emocionální kruh" který zahrnoval hypotalamus,
přední jádra thalamu, cingulární kůra, hipokampus a jejich spojení. Od té doby tyto struktury nazývají i fyziologové všude kolem Papets.
Pojem "viscerální mozek" navrhl P.D. McLean (1949), čímž označuje komplexní anatomicko-fyziologické sdružení, které se od roku 1952 začalo nazývat "limbický systém". Později se ukázalo, že limbický systém se podílí na výkonu různorodých funkcí a nyní je většina jeho části, včetně cingulárního a hipokampálního (parahipokampálního) gyri, obvykle kombinována do limbické oblasti, která má četná spojení se strukturami retikulární formace, tvořící s ní limbicko-retikulární komplex, který poskytuje široký kruh fyziologické a psychické procesy.
Aktuálně do limbický lalok Je zvykem připisovat prvky staré kůry (archiokortex) pokrývající gyrus dentatus a gyrus hippocampal; starověká kůra (paleokortex) předního hipokampu; stejně jako střední neboli intermediární kůra (mezokortex) gyru cingulate. Období "limbický systém" zahrnuje komponenty limbického laloku a přidružené struktury – entorinální (zabírající většinu parahipokampálního gyru) a septální oblasti, dále komplex amygdaly a mastoidní tělo (Duus P., 1995).
Mastoidní tělo spojuje struktury tohoto systému se středním mozkem a retikulární formací. Impulzy vznikající v limbickém systému mohou být přenášeny přes přední jádro thalamu do gyrus cingulate a do neokortexu po drahách tvořených asociativními vlákny. Impulzy pocházející z hypotalamu mohou dosáhnout orbitofrontálního kortexu a mediálního dorzálního jádra thalamu.
Četné přímé a zpětné vazby zajišťují propojení a vzájemnou závislost limbických struktur a mnoha útvarů diencefala a ústních částí trupu (nespecifická jádra thalamu, hypotalamu, putamenu, uzdičky, retikulární formace mozkového kmene), jakož i se subkortikálními jádry (globus pallidus , putamen, caudate nucleus) a s neokortexními mozkovými hemisférami, především s temporálním a frontálním lalokem.
Přes fylogenetické, morfologické a cytoarchitektonické odlišnosti jsou mnohé z uvedených struktur (limbická oblast, centrální a mediální struktury thalamu, hypotalamu, retikulární formace mozkového kmene) obvykle řazeny do tzv. limbicko-retikulární komplex, který působí jako zóna integrace více funkcí, zajišťující organizaci multimodálních, celostních reakcí těla na různé vlivy, což se projevuje zejména ve stresových situacích.
Struktury limbicko-retikulárního komplexu mají velký počet vstupy a výstupy, kterými procházejí uzavřené kruhy četných aferentních a eferentních spojení zajišťujících kombinované fungování formací zahrnutých v tomto komplexu a jejich interakce se všemi částmi mozku, včetně mozkové kůry.
Ve strukturách limbicko-retikulárního komplexu dochází ke sbližování senzitivních impulsů, které vznikají v intero- a exteroceptorech, včetně receptorových polí smyslových orgánů. Na tomto základě vzniká v limbicko-retikulárním komplexu primární syntéza informací o stavu vnitřního prostředí těla, i o vnějších faktorech prostředí působících na tělo a formují se elementární potřeby, biologické motivace a doprovodné emoce.
Limbicko-retikulární komplex určuje stav emocionální sféry, podílí se na regulaci vegetativně-viscerálních vztahů zaměřených na udržení relativní stálosti vnitřního prostředí (homeostázy), jakož i na zásobování energií a korelaci pohybových aktů. Na jeho stavu závisí úroveň vědomí, možnost automatizovaných pohybů, aktivita motorických a mentálních funkcí, řeč, pozornost, schopnost navigace, paměť, změna bdělosti a spánku.
Poškození struktur limbicko-retikulárního komplexu může být doprovázeno řadou klinických příznaků: výrazné změny v emoční sféře trvalého a záchvatovitého charakteru, anorexie nebo bulimie, sexuální poruchy, poruchy paměti, zejména známky Korsakoffova syndromu, kdy pacient ztrácí schopnost pamatovat si aktuální události (aktuální události jsou uchovávány v paměti po dobu ne delší než 2 minuty), vegetativně-endokrinní poruchy, poruchy spánku, psychosenzorické poruchy ve formě iluzí a halucinací, změny vědomí, klinické projevy akinetického mutismu, epileptické záchvaty.
Dosud bylo provedeno velké množství studií zaměřených na studium morfologie, anatomických souvislostí, funkce limbické oblasti a dalších struktur zahrnutých do limbicko-retikulárního komplexu, nicméně fyziologie a rysy klinického obrazu jeho poškození stále potřebují vyjasnění dnes. Většina informací o jeho funkci je zejména funkce parahipokampální oblasti, získané při pokusech na zvířatech metody dráždění, exstirpace nebo stereotaxe. Získané tímto způsobem výsledky vyžadují opatrnost při jejich extrapolaci na člověka. Zvláštní význam mají klinická pozorování pacientů s lézemi mediobazálních oblastí mozkové hemisféry.
V 50-60 letech XX století. V průběhu rozvoje psychochirurgie se objevily zprávy o léčbě pacientů s nevyléčitelnými duševními poruchami a syndromem chronické bolesti bilaterální cingulotomií (disekce gyru cingulate), přičemž obvykle byla zaznamenána regrese úzkosti, obsedantních stavů, psychomotorické agitace, bolestivých syndromů, které byla uznána jako důkaz účasti cingulárního gyru na utváření emocí a bolesti. Zároveň bisingulotomie vedla k hlubokým osobním poruchám, dezorientaci, snížení kritičnosti vlastního stavu a euforii.
Analýza 80 ověřených klinických pozorování hipokampálních lézí v Neurochirurgickém institutu Ruské akademie lékařských věd je uvedena v monografii N.N. Bragina (1974). Autor dochází k závěru, že temporální mediobazální syndrom zahrnuje viscerovegetativní, motorické a psychické poruchy, projevující se obvykle komplexně. Veškerá rozmanitost klinických projevů N.N. Bragin ji redukuje na dvě hlavní multifaktoriální varianty patologie s převahou „dráždivých“ a „inhibičních“ jevů.
První z nich zahrnuje emoční poruchy provázené motorickým neklidem (zvýšená vzrušivost, upovídanost, nervozita, pocit vnitřní úzkosti), záchvaty strachu, vitální melancholie, různé viscerovegetativní poruchy (změny pulsu, dýchání, gastrointestinální poruchy, zvýšená teplota, zvýšená pocení atd.). Tito pacienti na pozadí neustálého motorického neklidu často zažívali záchvaty motorické excitability.
nia. EEG této skupiny pacientů bylo charakterizováno mírnými mozkovými změnami směrem k integraci (rychlý a vyhrocený alfa rytmus, difuzní beta oscilace). Opakovaná aferentní stimulace vyvolala zřetelné EEG reakce, které na rozdíl od normálních nevymizely, když byly podněty opakovaně prezentovány.
Druhá („inhibiční“) verze mediobazálního syndromu je charakterizována emočními poruchami ve formě deprese s motorickou retardací (potlačená pozadí nálady, ochuzení a zpomalení tempa duševních procesů, změny v motorice, připomínající akineticko-rigidní syndrom Viscerovegetativní paroxysmy zaznamenané v první skupině jsou méně typické EEG pacientů v této skupině bylo charakterizováno celkovými mozkovými změnami, projevujícími se převahou pomalých forem aktivity (nepravidelný, pomalý alfa rytmus, skupiny oscilací theta, difúzní delta vlny Pozornost byla věnována prudkému poklesu reaktivity EEG.
Mezi těmito dvěma extrémy byly i střední s přechodnými a smíšenými kombinacemi jednotlivých symptomů. Některé z nich se tedy vyznačují poměrně slabými známkami agitované deprese se zvýšenou motorickou aktivitou a únavou, s převahou senestopatických vjemů, podezíravosti, dosahující u některých pacientů až paranoidních stavů a hypochondrických bludů. Další střední skupina se vyznačovala extrémní intenzitou depresivních symptomů na pozadí ztuhlosti pacienta.
Tyto údaje nám umožňují hovořit o duálním (aktivačním a inhibičním) vlivu hippocampu a dalších struktur limbické oblasti na behaviorální reakce, emoce, charakteristiky duševního stavu a bioelektrickou aktivitu kůry. Komplexní klinické syndromy tohoto typu by v současnosti neměly být považovány za primární fokální. Spíše je třeba je posuzovat ve světle představ o víceúrovňovém systému organizace mozkové činnosti.
S.B. Buklina (1997) poskytl údaje z vyšetření 41 pacientů s arteriovenózními malformacemi v oblasti gyrus cingulate. Před operací u 38 pacientů vystoupily do popředí poruchy paměti a u pěti z nich byly známky Korsakova syndromu, u tří pacientů vznikl Korsakovův syndrom po operaci, přičemž závažnost nárůstu poruch paměti korelovala s tzv. stupně destrukce vlastního gyru cingula i s postižením patologického procesu přilehlých struktur corpus callosum, přičemž amnestický syndrom nezávisel na straně malformace a její lokalizaci podél dlouhého gyru cingulate.
Hlavními charakteristikami zjištěných amnestických syndromů byly poruchy reprodukce sluchově-verbálních podnětů, zhoršená selektivita stop v podobě inkluzí a kontaminací a nezachování významu při předávání příběhu. Většina pacientů měla sníženou kritičnost při hodnocení svého stavu. Autor zaznamenal podobnost těchto poruch s amnestickými defekty u pacientů s frontálními lézemi, což lze vysvětlit přítomností spojení mezi gyrus cingulate a frontálním lalokem.
Více běžné patologické procesy v limbické oblasti způsobují těžké poruchy autonomně-viscerálních funkcí.
Corpus callosum(corpus callosum)- největší komisura mezi mozkovými hemisférami. Jeho přední části, zejména koleno callosum
tělo (genu corporis callosi), spojují čelní laloky, střední části - kmen corpus callosum (truncus corporis callosi)- zajišťují spojení mezi temporálními a parietálními úseky hemisfér, zadními úseky, zejména splenium corpus callosum (splenium corporis callosi), spojují okcipitální laloky.
Léze corpus callosum jsou obvykle doprovázeny duševními poruchami pacienta. Zničení jeho přední části vede k rozvoji „frontální psychiky“ (spontaneita, porušení plánu akce, chování, kritika, charakteristika frontální kalous syndrom - akineze, amimie, spontánnost, astasie-abasie, apraxie, úchopové reflexy, demence). Oddělení spojení mezi parietálními laloky vede k perverzi porozumění "tělesné diagramy" A vzhled apraxie hlavně v levé ruce. Disociace spánkových laloků může se objevit porucha vnímání vnějšího prostředí, ztráta správné orientace v něm (amnestické poruchy, konfabulace, syndrom již viděného a tak dále.). Patologická ložiska v zadních částech corpus callosum jsou často charakterizována známkami zrakové agnozie.
14.6. ARCHITEKTONIKA MOZKOVÉ KŮRY
Struktura mozkové kůry je heterogenní. Méně složité ve struktuře, které vznikají na počátku procesu fylogeneze starověká kůra (archiokortex) a stará kůra (paleokortex), příbuzný většinou do limbického laloku mozek Většina mozkové kůry (95,6 %) díky jejímu pozdějšímu vzniku z hlediska fylogeneze je tzv. nová kůra (neokortex) a má mnohem složitější vícevrstvou strukturu, ale také heterogenní ve svých různých zónách.
Kvůli architektonika kůry je v určité souvislosti s její funkcí, Jeho studiu byla věnována velká pozornost. Jedním ze zakladatelů doktríny cytoarchitektoniky kůry byl V.A. Betz (1834-1894), který v roce 1874 poprvé popsal velké pyramidové buňky motorického kortexu (Betzovy buňky) a určil principy dělení mozkové kůry na hlavní oblasti. Následně mnoho badatelů významně přispělo k rozvoji nauky o struktuře kůry - A. Cambell, E. Smith, K. Brodmann, Oskar Vogt a Cecilia Vogt, S. Vogt). Velké úspěchy ve studiu kortikální architektoniky patří týmu Brain Institute Akademie lékařských věd (S.A. Sarkisov, N.I. Filimonov, E.P. Kononova aj.).
Hlavní typ struktury nové kůry (obr. 14.5), se kterou jsou všechny její úseky srovnávány, je kůra, sestávající ze 6 vrstev (homotypická kůra podle Brodmanna).
Vrstva I je molekulární neboli zonální, nejpovrchnější, chudá na buňky, její vlákna mají směr převážně rovnoběžný s povrchem kůry.
Vrstva II - vnější granulovaná. Skládá se z velkého počtu hustě umístěných malých granulárních nervových buněk.
Vrstva III - malé a střední pyramidy, nejširší. Skládá se z pyramidálních buněk, jejichž velikosti jsou nestejné, což umožňuje ve většině korových polí tuto vrstvu rozdělit na podvrstvy.
Vrstva IV - vnitřní granulovaná. Skládá se z hustě umístěných malých granulovaných buněk, kulatých a hranatý tvar. Tato vrstva je nejvariabilnější, in
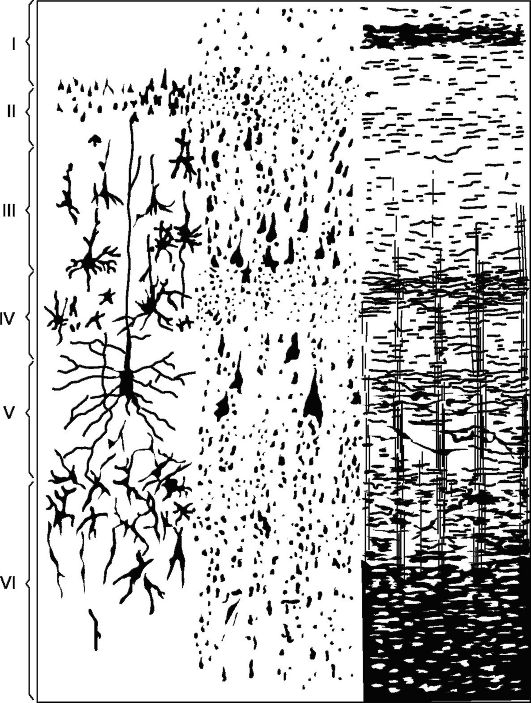
Rýže. 14.5.Cytoarchitektura a myeloarchitektura motorické zóny mozkové kůry.
Vlevo: I - molekulární vrstva; II - vnější zrnitá vrstva; III - vrstva malých a středních pyramid; IV - vnitřní zrnitá vrstva; V - vrstva velkých pyramid; VI - vrstva polymorfních buněk; vpravo - prvky myeloarchitektoniky.
V některých polích (například pole 17) se dělí na podvrstvy a někde se prudce ztenčuje a dokonce úplně mizí.
Vrstva V - velké pyramidy, neboli ganglion. Obsahuje velké pyramidální buňky. V některých oblastech mozku je vrstva rozdělena na podvrstvy, v motorické zóně se skládá ze tří podvrstev, z nichž prostřední obsahuje Betzovy obří pyramidální buňky, dosahující průměru 120 mikronů.
Vrstva VI - polymorfní buňky, nebo multiformní. Skládá se převážně z trojúhelníkových vřetenovitých buněk.
Struktura mozkové kůry má velké množství variací v důsledku změn tloušťky oddělené vrstvy ztenčení nebo vymizení nebo,
naopak ztluštěním a rozdělením na podvrstvy některých z nich (heterotypické zóny, dle Brodmanna).
Kůra každé mozkové hemisféry je rozdělena do několika oblastí: okcipitální, horní a dolní parietální, postcentrální, centrální gyri, precentrální, frontální, temporální, limbická, inzulární. Každý z nich podle vlastností je rozdělena do několika oborů, Každé pole má navíc své konvenční pořadové označení (obr. 14.6).
Studium architektoniky mozkové kůry spolu s fyziologickými, včetně elektrofyziologických, studiemi a klinickými pozorováními velkou měrou přispělo k vyřešení problematiky rozložení funkcí v kůře.
14.7. PROJEKČNÍ A ASOCIACE KORTÁLU
V procesu rozvíjení nauky o úloze mozkové kůry a jejích jednotlivých úseků při výkonu určitých funkcí se vyskytovala různá, někdy i protichůdná hlediska. Panoval tedy názor o přísně lokálním zastoupení v mozkové kůře všech lidských schopností a funkcí, až po ty nejsložitější, mentální (lokalizace, psychomorfologismus). Proti němu stál jiný názor o absolutní funkční ekvivalenci všech oblastí mozkové kůry. (ekvipotencialismus).
Významný příspěvek ke studiu lokalizace funkcí v mozkové kůře přinesl I.P. Pavlov (1848-1936). Identifikoval projekční zóny kůry (kortikální konce analyzátorů jednotlivé druhy citlivost) a asociativní zóny umístěné mezi nimi, studovali procesy inhibice a excitace v mozku, jejich vliv na funkční stav mozkové kůry. Rozdělení kůry na projekční a asociativní zóny přispívá k pochopení organizace práce mozkové kůry a ospravedlňuje se při řešení praktických problémů, zejména v aktuální diagnostice.
Projekční zóny poskytují především jednoduché specifické fyziologické úkony, především vnímání vjemů určité modality. Projekční dráhy, které se k nim přibližují, spojují tyto zóny s receptorovými územími na periferii, které jsou s nimi ve funkční korespondenci. Příklady projekčních kortikálních zón jsou oblast zadního centrálního gyru (zóna běžné typy citlivost) nebo oblast kalkarinového sulku umístěného na mediální straně okcipitálního laloku (projektivní zraková oblast).
Asociační zóny kůra nemá přímé spojení s periferií. Jsou umístěny mezi projekčními zónami a mají četná asociativní spojení s těmito projekčními zónami as jinými asociativními zónami. Funkcí asociativních zón je provádět vyšší analýzu a syntézu mnoha elementárních i složitějších komponent. Zde jsou v podstatě chápány informace vstupující do mozku a tvoří se myšlenky a koncepty.
G.I. Polyakov v roce 1969 na základě srovnání architektonické architektury mozkové kůry lidí a některých zvířat stanovil, že asociativní
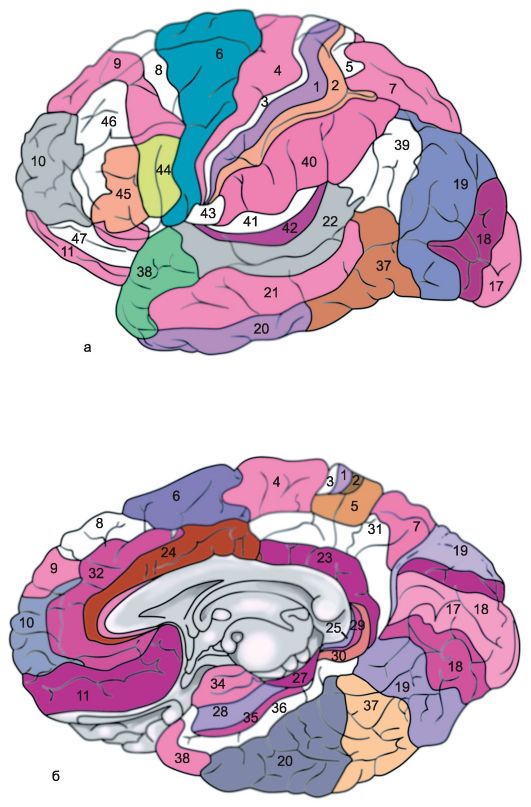
Rýže. 14.6.Architektonická pole mozkové kůry [podle Brodmanna]. a - vnější povrch; b - mediální plocha.
zóny v mozkové kůře člověka tvoří 50 %, v kůře vyšších (antropoidních) lidoopů - 20 %, u nižších lidoopů stejný údaj 10 % (obr. 14.7). Mezi asociačními zónami kůry lidský mozek, stejný autor navrhl izolovat sekundární a terciární obory. Sekundární asociativní pole sousedí s projekčními poli. Provádějí analýzu a syntézu elementárních vjemů, která si stále zachovává specifické zaměření.
Terciární asociační obory se nacházejí převážně mezi sekundárními a jsou zónami překrytí sousedních území. Souvisí především s analytickou činností mozkové kůry, zajišťující vyšší mentální funkce charakteristické pro člověka v jeho nejsložitějších intelektuálních a řečových projevech. Funkční vyspělost terciárního as-
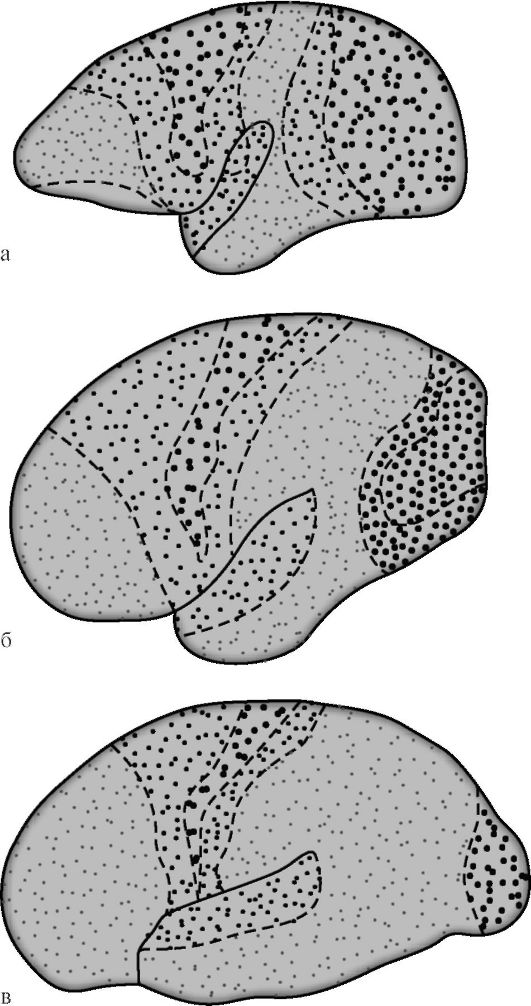
Rýže. 14.7. Diferenciace projekčních a asociačních zón mozkové kůry během evoluce primátů [podle G.I. Polyakov]. a - mozek nižší opice; b - mozek velké opice; c - lidský mozek. Velké tečky označují projekční zóny, malé tečky označují asociativní zóny. U nižších opic zabírají asociační zóny 10 % plochy kůry, u vyšších opic - 20 %, u lidí - 50 %.
sociálních polí mozkové kůry se vyskytuje nejpozději a to pouze v příznivém sociálním prostředí. Na rozdíl od jiných korových polí se terciální pole pravé a levé hemisféry vyznačují výrazným funkční asymetrie.
14.8. AKTUÁLNÍ DIAGNOSTIKA LÉZÍ MOZKOVÉ KŮRY
14.8.1. Projevy poškození projekčních zón mozkové kůry
V kůře každé mozkové hemisféry za centrálním gyrusem je 6 projekčních zón.
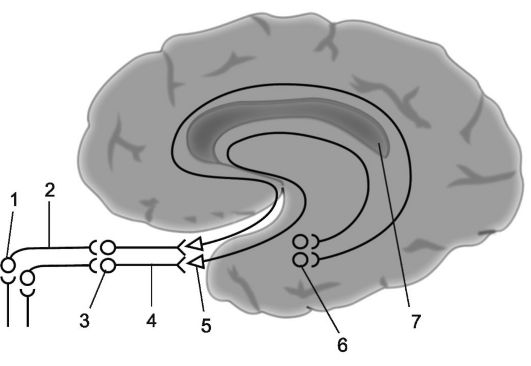
1. V přední části parietálního laloku, v oblasti zadního centrálního gyru (cytoarchitektonická pole 1, 2, 3) nachází se projekční zóna obecných typů citlivosti(obr. 14.4). Zde umístěné oblasti kůry přijímají citlivé impulsy přicházející po projekčních drahách obecných typů citlivosti z receptorového aparátu opačné poloviny těla. Čím vyšší je úsek této projekční zóny kůry, tím níže umístěné části protilehlé poloviny těla má projekční spoje. Části těla, které mají extenzivní příjem (jazyk, palmární povrch ruky), odpovídají neadekvátně velkým částem plochy projekčních zón, zatímco ostatní části těla (proximální končetiny, trup) mají malou plochu kortikální reprezentace.
Podráždění patologickým procesem kortikální zóny obecných typů citlivosti vede k záchvatu parestezie v částech těla odpovídajících podrážděným oblastem mozkové kůry (senzitivní Jacksonův záchvat), který se může vyvinout v sekundární generalizovaný paroxysmus. Poškození kortikálního konce analyzátoru obecných typů citlivosti může způsobit rozvoj hypalgezie nebo anestezie v odpovídající oblasti opačné poloviny těla, zatímco oblast hypoestezie nebo anestezie může být vertikální oběhové nebo radikulární segmentový typ. V prvním případě se porucha citlivosti projevuje na straně opačné k patologickému ohnisku v oblasti rtů, palce nebo v distální části končetiny s kruhovým okrajem, někdy jako ponožka nebo rukavice. Ve druhém případě má zóna zhoršené citlivosti tvar proužku a nachází se podél vnitřního nebo vnějšího okraje paže nebo nohy; to je vysvětleno skutečností, že vnitřní strana končetin je znázorněna vpředu a vnější strana - v zadních částech projekční zóny analyzátoru obecných typů citlivosti.
2. Zóna vizuální projekce nachází se v kůře mediálního povrchu okcipitálního laloku v oblasti kalkarinového žlábku (pole 17). V tomto poli je IV (vnitřní granulární) vrstva kortexu rozdělena na dvě podvrstvy svazkem myelinových vláken. Jednotlivé oblasti pole 17 přijímají impulsy z určitých částí homonymních polovin sítnice obou očí; v tomto případě zasahují impulsy přicházející ze spodních částí homonymních polovin sítnic
kůra dolního rtu calcarine sulcus a impulsy přicházející z horních částí sítnic směřují do kůry jeho horního rtu.
Poškození zóny zrakové projekce patologickým procesem vede ke vzniku kvadrantové nebo úplné homonymní hemianopsie na opačné straně patologického ohniska. Oboustranné poškození korových oblastí 17 nebo k nim vedoucích projekčních zrakových drah může vést k úplné slepotě. Podráždění kortexu zóny zrakové projekce může způsobit vzhled zrakových halucinací ve formě fotopsie v odpovídajících částech opačných polovin zorných polí.
3. Zóna sluchové projekce nachází se v kortexu Heschlova gyri na spodní ret laterální (Sylvian) trhlina (oblasti 41 a 42), které jsou ve skutečnosti součástí horního temporálního gyru. Podráždění této zóny kůry může způsobit výskyt sluchových halucinací (útoky pocitů hluku, zvonění, pískání, bzučení atd.). Zničení zóny sluchové projekce na jedné straně může způsobit mírné snížení sluchu v obou uších, v ve větší míře k opaku patologického zaměření.
4 a 5. Čichové a chuťové projekční zóny jsou na mediální ploše klenutého gyru (limbická oblast) mozku. První z nich se nachází v parahipokampálním gyru (pole 28). Projekční zóna chuti je obvykle lokalizována v operkulárním kortexu (oblast 43). Podráždění projekčních zón čichu a chuti může způsobit jejich zvrácení nebo vést k rozvoji odpovídajících čichových a chuťových halucinací. Jednostranná ztráta funkce projekčních zón čichu a chuti může způsobit mírné snížení čichu a chuti na obou stranách, resp. Oboustranná destrukce kortikálních konců stejných analyzátorů se projevuje absencí pachu a chuti na obou stranách, resp.
6. Vestibulární projekční zóna. Jeho lokalizace nebyla specifikována. Přitom je známo, že vestibulární aparát má četné anatomické a funkční souvislosti. Je možné, že lokalizace zastoupení vestibulárního systému v kortexu není dosud objasněna, protože je polyfokální. N.S. Blagoveshchenskaya (1981) se domnívá, že v mozkové kůře jsou vestibulární projekční zóny reprezentovány několika vzájemně interagujícími anatomickými a funkčními komplexy, které se nacházejí v poli 8, na křižovatce frontálních, temporálních a parietálních laloků a v oblasti centrálního gyru a předpokládá se, že každá z těchto oblastí kůry plní své vlastní funkce. Pole 8 je libovolné centrum pohledu, jeho dráždění způsobuje otočení pohledu opačným směrem k patologickému ohnisku, změny rytmu a charakteru experimentálního nystagmu, zvláště brzy po epileptickém záchvatu. V kůře spánkového laloku jsou útvary, jejichž podráždění způsobuje závratě, které se projevují zejména epilepsií spánkového laloku; poškození oblastí zastoupení vestibulárních struktur v kůře centrálního gyru ovlivňuje stav tonusu příčně pruhovaných svalů. Klinická pozorování naznačují, že jaderně-kortikální vestibulární dráhy procházejí částečnou dekusací.
Je třeba zdůraznit, že známky podráždění uvedených projekčních zón mohou být projevem aury epileptického záchvatu odpovídajícího charakteru.
I.P. Pavlov považoval za projekční zónu gyru precentrální, který ovlivňuje motorické funkce a svalový tonus převážně opačné poloviny těla, se kterou je spojen především kortikonukleární a kortikospinální (pyramidální) dráhou. takzvaný analyzátor motoru. Tato zóna zabírá Především pole 4, na které se v převrácené podobě promítá především protilehlá polovina těla. Toto pole obsahuje většinu obřích pyramidálních buněk (Betzovy buňky), jejichž axony tvoří 2-2,5 % všech vláken pyramidálního traktu, a také střední a malé pyramidové buňky, které spolu s axony téhož buňky umístěné v oblasti sousedící s polem 4 a rozsáhlejším polem 6, se podílejí na realizaci monosynaptických a polysynaptických kortikosvalových spojení. Monosynaptická spojení zajišťují především rychlé a přesné cílené akce v závislosti na kontrakcích jednotlivých pruhovaných svalů.
Poškození dolní motorické oblasti obvykle vede k rozvoji na opačné straně brachiofaciální (humerofaciální) syndrom nebo linguofaciálně-brachiální syndrom, které jsou často pozorovány u pacientů s cévní mozkovou příhodou v povodí střední mozkové tepny, s kombinovanou parézou svalů obličeje, jazyka a paže, především ramene, centrálního typu.
Podráždění kůry motorické zóny (pole 4 a 6) vede ke vzniku spasmů ve svalech nebo svalových skupinách promítaných do této zóny. Častěji se jedná o lokální záchvaty Jacksonského typu epilepsie, které se mohou přeměnit v sekundární generalizovaný epileptický záchvat.
14.8.2. Projevy poškození asociativních polí mozkové kůry
Mezi projekčními zónami kůry jsou asociativní pole. Impulsy dostávají především z buněk projekčních zón kůry. V asociativních polích dochází k analýze a syntéze informací, které prošly primárním zpracováním v projekčních polích. Asociativní zóny kůry horního parietálního laloku poskytují syntézu elementárních vjemů, proto se zde vytvářejí komplexní typy citlivosti, jako je smysl pro lokalizaci, smysl pro váhu, dvourozměrný prostorový smysl a také komplexní kinestetické vjemy.
V oblasti interparietálního sulku se nachází asociativní zóna, která zajišťuje syntézu vjemů vycházejících z částí vlastního těla. Poškození této oblasti kůry vede k autotopagnózie, těch. k nesprávnému rozpoznání nebo neznalosti částí vlastního těla, nebo k pseudomelie - pocit, že máte navíc ruku nebo nohu, stejně jako anosognosie - nedostatečné povědomí o fyzické vadě, která vznikla v souvislosti s onemocněním (například ochrnutí nebo paréza končetiny). Obvykle se všechny typy autotopagnosie a anosognosie vyskytují, když je patologický proces umístěn vpravo.
Poškození dolního parietálního lalůčku se může projevit poruchou syntézy elementárních vjemů nebo neschopností porovnat syntetizované komplexní vjemy s tím, co bylo kdysi ve vjemu podobné.
stejným způsobem, na základě jehož výsledků dochází k uznání“ (V.M. Bekhterev). To se projevuje porušením dvourozměrného prostorového smyslu (grafestézie) a trojrozměrného prostorového smyslu (stereognóza) - astereognóze.
V případě poškození premotorických zón frontálního laloku (pole 6, 8, 44) obvykle dochází k frontální ataxii, při které dochází k syntéze aferentních impulsů (kinestetická aferentace), signalizujících měnící se polohu částí těla v prostoru při pohybech. , je narušen.
Při poruše funkce kůry předních částí čelního laloku, která má spojení s opačnou hemisférou mozečku (frontopontinně-cerebelární spojení), dochází k poruchám statokinetiky na straně protilehlé patologickému ohnisku. (frontální ataxie). Obzvláště zřetelná jsou porušení pozdně se vyvíjejících forem statokinetiky – vzpřímený stoj a vzpřímená chůze. V důsledku toho pacient pociťuje nejistotu a nejistou chůzi. Při chůzi se tělo naklání dozadu (Hennerův příznak) položí nohy v přímce (liščí procházka) někdy při chůzi dochází k „opletení“ nohou. U některých pacientů s poškozením předních částí čelních laloků se vyvine zvláštní jev: při absenci paralýzy a parézy a schopnosti plně pohybovat nohama nemohou pacienti stát (astasia) a chodit (abázie).
Poškození asociativních zón kortexu je často charakterizováno rozvojem klinických projevů poruch vyšších psychických funkcí (viz kap. 15).
Obecný přehled stavby mozkových hemisfér
Mozkové hemisféry jsou nejhmotnější částí mozku. Pokrývají mozeček a mozkový kmen. Mozkové hemisféry tvoří přibližně 78 % celkové mozkové hmoty. Při ontogenetickém vývoji organismu se z mozkového váčku nervové trubice vyvíjejí mozkové hemisféry, proto se této části mozku také říká tzv. telencephalon.
Mozkové hemisféry jsou rozděleny podél střední čáry hlubokou vertikální trhlinou na pravou a levou hemisféru.
V hloubce střední části jsou obě hemisféry navzájem spojeny velkou komisurou - corpus callosum. Každá hemisféra má laloky; frontální, parietální, temporální, okcipitální a insula.
Laloky mozkových hemisfér jsou od sebe odděleny hlubokými rýhami. Nejdůležitější jsou tři hluboké rýhy: centrální (rolandský) oddělující frontální lalok od temenního, laterální (Sylvian) oddělující spánkový lalok od temenního, parietookcipitální oddělující temenní lalok od týlního na vnitřní ploše polokouli.
Každá hemisféra má superolaterální (konvexní), spodní a vnitřní povrch.
Každý lalok hemisféry má mozkové závity oddělené od sebe žlábky. Horní část hemisféry je pokryta kůrou ~ tenkou vrstvou šedé hmoty, která se skládá z nervových buněk.
Mozková kůra je z evolučního hlediska nejmladším útvarem centrálního nervového systému. U lidí dosahuje svého nejvyššího rozvoje. Mozková kůra má velký význam při regulaci vitálních funkcí těla, při realizaci komplexních forem chování a rozvoji neuropsychických funkcí.
Pod kůrou je bílá hmota hemisfér, skládá se z výběžků nervových buněk - vodičů. V důsledku tvorby mozkových konvolucí se výrazně zvětšuje celkový povrch mozkové kůry. Celková plocha mozkové kůry je 1200 cm2, přičemž 2/3 jejího povrchu jsou umístěny v hloubce rýh a 1/3 na viditelném povrchu hemisfér. Každý lalok mozku má jiný funkční význam.
a – superolaterální plocha: 1 – gyrus frontalis inferior; 2 – střední frontální gyrus; 3 – gyrus frontalis superior; 4 – přední centrální gyrus; 5 – centrální (rolandský) sulcus; 6 – zadní centrální gyrus; 7 – horní parietální lalok; 8 – dolní parietální lalůček; 9 – supramarginální rýha; 10 – hranatá (hranatá) drážka; 11 – sulcus parieto-okcipitální; 12 – gyrus temporalis inferior;
13 – střední temporální gyrus; 14 – gyrus temporalis superior; 15 – laterální (Sylvian) puklina;
b – vnitřní plocha: 1 paracentrální lalůček; 2 – středová drážka; 3 – gyrus cingulate: 4 – corpus callosum; 5 – sulcus parieto-okcipitální; 6 – klín; 7 – kalkarinová drážka; 8 – gyrus lingulární; 9 – gyrus hippocampal (gyrus parahipocampal).
Stavba čelního laloku
Čelní lalok zaujímá přední části hemisfér. Od parietálního laloku je oddělen centrálním sulkem a od temporálního laloku laterálním sulkem. Frontální lalok má čtyři gyry: jeden vertikální - precentrální a tři horizontální - horní, střední a dolní frontální gyri. Závity jsou od sebe odděleny drážkami.
Na spodní ploše čelních laloků se rozlišují přímé a orbitální gyri. Gyrus recta leží mezi vnitřním okrajem hemisféry, olfactorius sulcus a vnějším okrajem hemisféry.
V hlubinách čichového sulku leží čichový bulbus a čichový trakt.
Lidský čelní lalok tvoří 25–28 % mozkové kůry; průměrná hmotnost čelního laloku je 450 g.
Funkce čelních laloků je spojena s organizací dobrovolných pohybů, motorickými mechanismy řeči, regulací složitých forem chování a procesů myšlení. Několik funkčně důležitých center je soustředěno v konvolucích frontálního laloku. Přední centrální gyrus je „reprezentací“ primární motorické zóny s přesně definovanou projekcí částí těla. Obličej je „umístěn“ v dolní třetině gyru, ruka je ve střední třetině, noha je v horní třetině. Trup je zastoupen v zadních částech horního frontálního gyru. Člověk je tedy promítán v předním centrálním gyru hlavou dolů a hlavou dolů.
Přední centrální gyrus spolu s přilehlým zadním a částmi frontálního gyru hraje velmi důležitou funkční roli. Je centrem dobrovolných hnutí. Hluboko v kůře centrálního gyru začíná hlavní motorická dráha od takzvaných pyramidových buněk - centrálního motorického neuronu - pyramidální, kortikospinální, cesta. Periferní procesy motorických neuronů opouštějí kůru, shromažďují se do jediného mocného svazku, procházejí centrální bílou hmotou hemisfér a vstupují do mozkového kmene přes vnitřní pouzdro; na konci mozkového kmene částečně dekusují (přecházejí z jedné strany na druhou) a poté sestupují do míchy. Tyto procesy končí v šedé hmotě míchy. Tam se dostávají do kontaktu s periferním motorickým neuronem a přenášejí do něj impulsy z centrálního motorického neuronu. Impulzy dobrovolného pohybu jsou přenášeny po pyramidální dráze.
V zadních částech horního frontálního gyru je také extrapyramidové centrum kůry, anatomicky i funkčně úzce spojeny s útvary tzv. extrapyramidového systému. Extrapyramidový systém je motorický systém, který pomáhá při dobrovolném pohybu. Jedná se o systém „poskytování“ dobrovolných pohybů. Fylogeneticky starší extrapyramidový systém u lidí zajišťuje automatickou regulaci „naučených“ motorických aktů, udržení celkového svalového tonu, připravenost periferního motorického systému k provádění pohybů a redistribuci svalového tonu při pohybech. Kromě toho se podílí na udržení normálního držení těla.
Oblasti motorické kůry jsou lokalizovány především v precentrálním gyru (oblasti 4 a 6) a paracentrálním laloku na mediální ploše hemisféry. Existují primární a sekundární oblasti - pole 4 a 6. Tato pole jsou motorická, ale podle svých charakteristik se podle výzkumu Brain Institute liší. V primární motorické kůře(pole 4) jsou neurony inervující motorické neurony svalů obličeje, trupu a končetin.
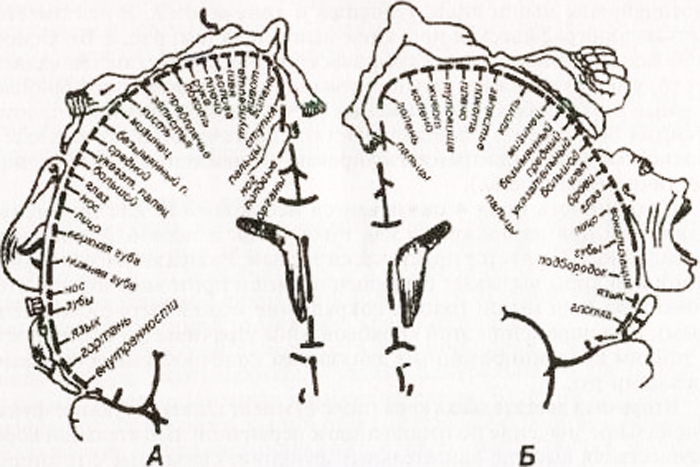
Schéma autototopické projekce obecné citlivosti a motorických funkcí v mozkové kůře (podle W. Penfielda):
A – kortikální projekce obecné citlivosti; B – kortikální projekce motorického systému. Relativní velikosti orgánů odrážejí oblast mozkové kůry, ze které mohou být vyvolány odpovídající pocity a pohyby
Má jasnou topografickou projekci svalů těla. Hlavním vzorem topografického zobrazení je, že regulace činnosti svalů, které zajišťují nejpřesnější a nejrozmanitější pohyby (řeč, psaní, mimika), vyžaduje účast velkých oblastí motorického kortexu. Pole 4 je zcela obsazeno středy izolovaných pohybů, pole 6 je obsazeno pouze částečně (podpole 6a).
Ukazuje se, že zachování pole 4 je nezbytné pro získání pohybů, když jsou stimulovány pole 4 i pole 6. U novorozence je pole 4 téměř zralé. Podrážděním primární motorické kůry dochází ke kontrakci svalů opačné strany těla (u svalů hlavy může být kontrakce oboustranná). Při poškození této kortikální zóny se ztrácí schopnost provádět jemné koordinované pohyby končetin a zejména prstů.
Sekundární motorická kůra(pole 6) má dominantní funkční význam ve vztahu k primární motorické kůře, která vykonává vyšší motorické funkce spojené s plánováním a koordinací volních pohybů. Zde je v největší míře zaznamenán pozvolna narůstající zápor. potenciál připravenosti, vyskytující se přibližně 1 s před začátkem pohybu. Kůra oblasti 6 přijímá většinu impulsů z bazálních ganglií a mozečku a podílí se na zaznamenávání informací o komplexních pohybech.
Podráždění kortexu oblasti 6 způsobuje složité koordinované pohyby, například otáčení hlavy, očí a trupu opačným směrem, kooperativní kontrakce flexorů nebo extenzorů na opačné straně. Premotorická kůra obsahuje motorická centra spojená s sociální funkcečlověk: centrum psané řeči v zadní části středního frontálního gyru (pole 6), centrum motorické řeči Broca v zadní části dolního frontálního gyru (pole 44), zajišťující řeč, stejně jako hudebně motorické centrum (pole 45), poskytující tón řeči, schopnost zpívat . Spodní část pole b (podpole bor), umístěné v oblasti pneumatiky, reaguje na elektrický proud rytmickými žvýkacími pohyby. Neurony motorického kortexu přijímají aferentní vstupy přes thalamus ze svalových, kloubních a kožních receptorů, z bazálních ganglií a cerebellum. Hlavním eferentním výstupem motorického kortexu do kmene a motorických center páteře jsou pyramidové buňky vrstvy V.
V zadní části středního frontálního gyru se nachází frontální okohybné centrum, které řídí současnou, současnou rotaci hlavy a očí (střed rotace hlavy a očí v opačném směru). Podráždění tohoto centra způsobuje otáčení hlavy a očí opačným směrem. Funkce tohoto centra má velký význam při realizaci tzv. orientačních reflexů (neboli reflexů „co to je?“), které jsou velmi důležité pro zachování života zvířat.
Frontální kůra mozkových hemisfér se také aktivně podílí na utváření myšlení, organizaci cílevědomých činností a dlouhodobém plánování.
Stavba parietálního laloku
Parietální lalok zaujímá horní boční plochy hemisféry. Z čelního laloku je temenní lalok omezen zepředu a do strany centrálním žlábkem, ze spánkového laloku níže - laterální rýhou, z týlního - pomyslnou linií probíhající od horního okraje parietookcipitálního sulcus ke spodnímu okraji hemisféry.
Na superolaterálním povrchu parietálního laloku jsou tři gyry: jeden vertikální - zadní centrální a dva horizontální - horní parietální a dolní parietální. Část gyrus parietalis inferior, která obepíná zadní část sulcus lateralis, se nazývá supramarginální (supramarginální) oblast, část obklopující gyrus temporalis superior je oblast nodální (úhlová).
Temenní lalok, stejně jako čelní lalok, tvoří významnou část mozkových hemisfér. Z fylogenetického hlediska se rozlišuje staré oddělení- zadní centrální gyrus, nový - gyrus parietalis superior a novější - gyrus parietalis inferior.
Funkce parietálního laloku je spojena s vnímáním a analýzou smyslových podnětů a prostorovou orientací. Několik funkčních center je soustředěno v gyri parietálního laloku.
V zadním centrálním gyru se centra citlivosti promítají s projekcí těla podobnou té v předním centrálním gyru. Obličej se promítá do dolní třetiny gyrusu, paže a trup se promítají do střední třetiny a noha se promítá do horní třetiny. V horním parietálním gyru jsou zodpovědná centra komplexní druhy hluboká citlivost: svalově-kloubní, dvourozměrný prostorový smysl, smysl pro hmotnost a rozsah pohybu, smysl pro rozpoznávání předmětů hmatem.
Za horními částmi zadního centrálního gyru je umístěno centrum, které poskytuje schopnost rozpoznat vlastní tělo, jeho části, jejich proporce a relativní polohy (pole 7).
Pole 1, 2, 3 postcentrální oblasti tvoří hlavní kortikální jádro kožního analyzátoru. Spolu s polem 1 je pole 3 primární a pole 2 sekundární projekční zóna kůže. analyzátor. Postcentrální oblast je propojena eferentními vlákny s podkorovými a kmenovými útvary, s precentrálními a dalšími oblastmi mozkové kůry. Kortikální část citlivého analyzátoru je tedy lokalizována v parietálním laloku.
Primární smyslové oblasti - jsou to oblasti smyslové kůry, jejichž podráždění nebo destrukce způsobuje jasné a trvalé změny v citlivosti těla (jádra analyzátoru, podle I.P. Pavlova). Skládají se převážně z unimodálních neuronů a tvoří vjemy stejné kvality. V primárních senzorických zónách je obvykle zřetelné prostorové (topografické) znázornění částí těla a jejich receptorových polí.
Kolem primárních senzorických oblastí jsou méně lokalizované sekundární smyslové oblasti jejichž neurony reagují na působení více podnětů, tzn. jsou multimodální.
Nejdůležitější senzorickou oblastí je parietální kůra postcentrálního gyru a odpovídající část paracentrálního laloku na mediální ploše hemisfér, která je označena jako somatosenzorická oblast I. Zde dochází k projekci kožní citlivosti na opačnou stranu těla z hmatových, bolestivých, teplotních receptorů, interoceptivní citlivosti a citlivosti pohybového aparátu – ze svalových, kloubních, šlachových receptorů.
Kromě somatosenzorické oblasti I se rozlišují menší velikosti somatosenzorický oblast II, nachází se na hranici průsečíku centrálního sulku s horním okrajem temporálního laloku, v hloubce laterálního sulku. Míra lokalizace částí těla je zde méně výrazná.
Nachází se ve spodním parietálním laloku praxi center. Praxis se týká účelových pohybů, které se zautomatizovaly v procesu opakování a cvičení, které se vyvíjejí v procesu učení a neustálého cvičení po určitou dobu. individuální život. Chůze, jídlo, oblékání, mechanický prvek psaní, různé druhy pracovní činnost(například pohyby řidiče při jízdě, sečení atd.) jsou praxí. Praxe je nejvyšším projevem motorické funkce vlastní člověku. Provádí se jako výsledek kombinované činnosti různých oblastí mozkové kůry.
V dolních částech předního a zadního centrálního gyru je centrum interoceptivního analyzátoru impulsů vnitřní orgány a plavidla. Střed má úzké spojení se subkortikálními vegetativními formacemi.
Stavba spánkového laloku
Spánkový lalok zaujímá inferolaterální povrch hemisfér. Z frontálního a parietálního laloku je temporální lalok omezen laterálním sulcusem. Na superolaterálním povrchu spánkového laloku jsou tři gyry: horní, střední a dolní.
Gyrus temporalis superior se nachází mezi Sylvian a superior temporal fisurs, prostřední je mezi horní a dolní temporální rýhou a dolní je mezi sulcus temporal inferior a transversální medulární fisurou. Na spodní ploše spánkového laloku se rozlišuje gyrus temporalis inferior, gyrus occipitotemporalis lateralis a gyri hippocampal (noha mořského koníka).
Funkce spánkového laloku je spojena s vnímáním sluchových, chuťových, čichových vjemů, analýzou a syntézou zvuků řeči a paměťovými mechanismy. Hlavní funkční centrum horní laterální plochy spánkového laloku se nachází v horním temporálním gyru. Nachází se zde sluchové neboli gnostické centrum řeči (Wernickeovo centrum).
Dobře prozkoumanou zónou primární projekce je sluchová kůra (pole 41, 42, 52), která se nachází hluboko v laterálním sulku (kůra Heschlových příčných temporálních gyri). Projekční kůra spánkového laloku zahrnuje také vestibulární analyzátor centrum● v horním a středním temporálním gyru (pole 20 a 21).
Oblast čichové projekce se nachází v gyru hippocampu, zejména v jeho přední části (tzv. uncus). Vedle zón čichové projekce jsou zóny chuťové.
Spánkové laloky hrají důležitou roli při organizování složitých mentálních procesů, zejména paměti.
Stavba týlního laloku
Týlní lalok zaujímá zadní části hemisfér. Na konvexním povrchu hemisféry nemá okcipitální lalok žádné ostré hranice oddělující jej od parietálního a temporálního laloku, s výjimkou horní části parieto-okcipitálního sulcus, která se nachází na vnitřním povrchu hemisféry a odděluje temenní lalok z týlního laloku. Drážky a konvoluce superolaterální plochy týlního laloku nejsou konstantní a mají proměnlivou strukturu. Na vnitřním povrchu týlního laloku je kalkarinní rýha, která odděluje cuneus (trojúhelníkový lalůček týlního laloku) od gyru lingualis a gyrus occipitotemporalis.
Funkce okcipitálního laloku je spojena s vnímáním a zpracováním vizuálních informací, organizací složitých procesů zrakového vnímání - V tomto případě se horní polovina sítnice promítá do oblasti klínu, přijímá světlo z dolních polí vidění; v oblasti lingulárního gyru se nachází dolní polovina sítnice oka, která vnímá světlo z horních zorných polí.
Nachází se v okcipitální kůře primární vizuální oblast(kortex části gyru sphenoidálního a lingválního lalůčku, oblast 17). Zde je aktuální zastoupení retinálních receptorů. Každý bod sítnice odpovídá své vlastní části zrakové kůry, zatímco zóna makuly má relativně velkou oblast zastoupení. V důsledku neúplného rozrušení zrakových drah jsou stejné poloviny sítnice promítány do vizuální oblasti každé hemisféry. Přítomnost retinální projekce v obou očích v každé hemisféře je základem binokulárního vidění. Poblíž pole 17 se nachází kůra sekundární zraková oblast(pole 18 a 19). Neurony těchto zón jsou multimodální a reagují nejen na světlo, ale také na hmatové a sluchové podněty. V této zrakové oblasti se syntetizují různé druhy citlivosti, vznikají složitější vizuální obrazy a provádí se jejich rozpoznání.
Insula, limbická kůra
Ostrov, nebo tzv. uzavřený lalůček, se nachází v hloubce laterálního sulku. Izolace je oddělena od sousedních sousedních částí kruhovou drážkou. Povrch insuly je rozdělen podélnou středovou rýhou na přední a zadní část. Na ostrově je promítán analyzátor chuti.
Limbická kůra. Na vnitřním povrchu hemisfér nad corpus callosum je gyrus cingulate. Tento gyrus prochází šíjí za corpus callosum do gyru poblíž mořského koníka - parahipokampálního gyru. Gyrus cingulární tvoří spolu s gyrus parahipokampální gyrus klenutý.
Vnitřní a spodní plochy hemisfér jsou spojeny do tzv. limbické (okrajové) kůry spolu s jádrem amygdaly ze skupiny subkortikálních jader, čichovým traktem a bulbem, oblastmi čelního, spánkového a temenního laloku mozkového. kůra, stejně jako subtuberkulární oblast a retikulární formace trupu. Limbická kůra je sjednocena do jediného funkčního systému – limbicko-retikulárního komplexu. Hlavní funkcí těchto částí mozku není ani tak poskytovat komunikaci s vnějším světem, ale regulovat tonus kůry, pohony a afektivní život. Regulují komplexní, mnohostranné funkce vnitřních orgánů a behaviorální reakce. Limbicko-retikulární komplex je nejdůležitější integrační systém těla. Limbický systém je také důležitý při formování motivace. Motivace (neboli vnitřní pohon) zahrnuje nejsložitější instinktivní a emocionální reakce(potravinové, obranné, sexuální). Limbický systém se také podílí na regulaci spánku a bdění.
Limbická kůra také plní důležitou funkci čichu. Čich – vnímání těch ve vzduchu chemické substance. Lidský čichový mozek zajišťuje čich, stejně jako organizaci složitých forem emocionálních a behaviorálních reakcí. Čichový mozek je součástí limbického systému.
Čichový mozek se skládá ze dvou částí – periferní a centrální. Periferní část představuje čichový nerv, čichové bulby a primární čichová centra. Centrální část zahrnuje gyrus mořského koníka - hippocampus, zubatý a klenutý gyr.
Čichový receptorový aparát se nachází v nosní sliznici. Prostřednictvím systému nervových vodičů jsou informace z receptorů přenášeny do kortikální části čichového analyzátoru.
Kortikální část čichového analyzátoru se nachází v gyru cingulate, gyrus mořského koníka a uncus mořského koníka, které dohromady tvoří uzavřenou prstencovitou oblast. Periferní část čichového analyzátoru je napojena na korové oblasti obou hemisfér.
Fyziologický mechanismus vnímání pachů čichovým analyzátorem není zcela jasný. Existují dvě hlavní hypotézy, s různé polohy vysvětlující podstatu tohoto procesu. Podle jedné hypotézy k interakci mezi molekulami odorantu a chemoreceptory dochází jako klíč a zámek, tzn. typ molekuly odpovídá speciálnímu receptoru. Další hypotéza je založena na předpokladu, že molekuly vonné látky mají určitou vibrační vlnu, na kterou jsou „naladěny“ čichové receptory. Molekuly, které mají podobné vibrace, musí mít společnou vlnu, a proto vydávají podobné pachy.
Termín „čichový mozek“ ve vztahu k lidské fyziologii je poněkud svévolný a neodhaluje plně jeho mnohostrannou a univerzální funkci. "Umístění" centrálního článku čichový mozek v mozkových hemisférách není náhodný a je výsledkem obrovské „informační“ role, kterou čich hrál v procesu evoluce při adaptaci na vnější prostředí a regulaci komplexních behaviorálních reakcí. Získávání potravy, výběr jedince opačného pohlaví, péče o potomstvo, celistvost teritoria, organizování skupinových společenstev v rámci druhu – všechny tyto každodenní funkce u mnoha zvířat probíhají za přímé účasti jemně vybudovaného čichového přijímacího systému a schopnost řady živočichů na základě toho vysílat do vnějšího prostředí jemné diferencované signály.informační signály jsou specifické pachové látky.
V lidském životě ztratil čich biologický informační význam, který měl u zvířat. Lidský čichový systém je navržen tak, aby vykonával úzkou, „svou vlastní“ funkci, a také pro jakési „nabíjení“ emocí. Síla vlivu pachů na emocionální sféru, skutečnost, že jsou nejdůležitějším „potravinovým substrátem emocí“, je dobře známá již od pradávna v historii lidstva.
Čich člověka se může lišit. Tyto variace jsou zpravidla nevýznamné, ale v některých případech může být ostrost čichu velmi vysoká (degustátoři v parfémovém průmyslu).
Protože čichový analyzátor hraje důležitou roli v regulaci emocí, je jeho centrální část klasifikována jako limbický systém, obrazně nazývaný „společný jmenovatel“ mnoha emočních a viscerosomatických reakcí těla.
Centrum analyzátoru chuti se nachází v bezprostřední blízkosti středu čichového analyzátoru, tzn. v háku a ammonově cornu, ale navíc v nejnižší části zadního centrálního gyru (oblast 43), stejně jako v insula. Stejně jako čichový analyzátor poskytuje centrum funkci projekce, ukládání a rozpoznávání chuťových obrazů.
Nachází se na hranici temporálního, okcipitálního a parietálního laloku centrum pro analýzu písemné řeči(pole 39), které je úzce spojeno s Wernickeho centrem spánkového laloku, s centrem zrakového analyzátoru týlního laloku a také s centry parietálního laloku. Čtenářské centrum poskytuje rozpoznávání a ukládání obrazů psaného jazyka.
Základní informace o prostředí a vnitřní prostředí těla, přijaté v senzorické kůře, je převedeno k dalšímu zpracování do asociativní kůry, načež je iniciováno (v případě potřeby) behaviorální reakce s povinnou účastí motorického kortexu.
Corpus callosum– obloukovitá tenká deska, fylogeneticky mladá, spojuje střední plochy obou hemisfér. Protáhlá střední část corpus callosum vzadu ztlušťuje a vpředu se obloukovitě ohýbá a ohýbá dolů. Corpus callosum spojuje fylogeneticky nejmladší části hemisfér a hraje důležitou roli při výměně informací mezi nimi.
Architektonika mozkové kůry
Studium strukturálních rysů struktury kůry se nazývá architektonika. Buňky mozkové kůry jsou méně specializované než neurony v jiných částech mozku; nicméně určité jejich skupiny jsou anatomicky a fyziologicky úzce spjaty s určitými specializovanými částmi mozku.
Mikroskopická struktura mozkové kůry je v různých částech odlišná. Tyto morfologické rozdíly v kortexu nám umožnily identifikovat samostatná kortikální cytoarchitektonická pole. Existuje několik možností klasifikace korových polí. Většina výzkumníků identifikuje 50 cytoarchitektonických polí, jejichž mikroskopická struktura je poměrně složitá.
Kůra se skládá ze 6 vrstev buněk a jejich vláken. Hlavní typ struktury kůry je šestivrstevný, ne všude je však jednotný. Existují oblasti kůry, kde je jedna z vrstev výrazně vyjádřena a druhá je vyjádřena slabě. V jiných oblastech kůry jsou některé vrstvy rozděleny na podvrstvy atd.
Bylo zjištěno, že oblasti kůry spojené se specifickou funkcí mají podobnou strukturu. Oblasti kůry, které jsou svým funkčním významem blízké u zvířat a lidí, mají určitou podobnost ve struktuře. Ty části mozku, které vykonávají čistě lidské funkce (řeč), jsou přítomny pouze v lidské kůře a chybí u zvířat, dokonce ani u opic.
Morfologická a funkční heterogenita mozkové kůry umožnila identifikovat centra zraku, sluchu, čichu atd., která mají svou specifickou lokalizaci. Je však nesprávné hovořit o kortikálním centru jako o přísně omezené skupině neuronů. Specializace oblastí kůry se formuje v procesu života. Na začátku dětství Funkční zóny kůry se navzájem překrývají, takže jejich hranice jsou nejasné a nezřetelné. Pouze v procesu učení, akumulace vlastní zkušenost praktickou činností dochází k postupnému soustřeďování funkčních zón do od sebe oddělených center.
Bílá hmota mozkových hemisfér se skládá z nervových vodičů. Podle anatomických a funkčních charakteristik se vlákna bílé hmoty dělí na asociativní, komisurální a projekční.

a – asociativní vlákna; b – komisurální vlákna; b-projekční vlákna.
Asociativní vlákna spojují různé oblasti kůry v rámci jedné hemisféry. Tato vlákna jsou krátká a dlouhá. Krátká vlákna mají obvykle obloukovitý tvar a spojují sousední gyri. Dlouhá vlákna spojují vzdálené oblasti kůry. Komisní vlákna se obvykle nazývají ta vlákna, která spojují topograficky identické oblasti pravé a levé hemisféry. Komisurální vlákna tvoří tři komisury: přední bílá komisura, fornixová komisura a corpus callosum. Přední bílá komisura spojuje čichové oblasti pravé a levé hemisféry. Fornixová komisura spojuje hipokampální gyri pravé a levé hemisféry. Převážná část komisurálních vláken prochází corpus callosum a spojuje symetrické oblasti obou hemisfér mozku.
Projektivní jsou ta vlákna, která spojují mozkové hemisféry se základními částmi mozku – kmenem a míchou. Projekční vlákna obsahují dráhy nesoucí aferentní (senzitivní) a eferentní (motorické) informace.
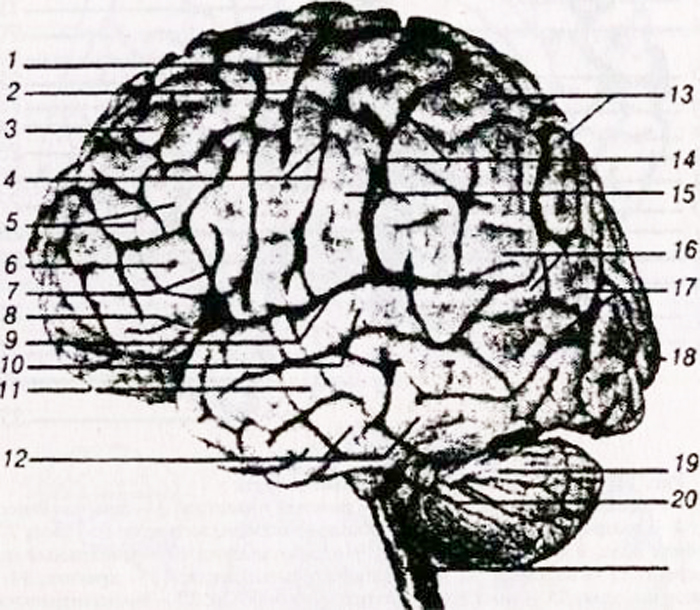
1 – precentrální gyrus; 2 – precentrální sulcus; 3 – gyrus frontalis superior; 4 – středová drážka; 5 – střední frontální gyrus;
b – gyrus frontální inferior; 7 – vzestupná větev laterálního sulcus;
8 – horizontální větev laterálního sulcus; 9 – zadní větev laterálního sulcus; 10 – gyrus temporalis superior; 11 – střední temporální gyrus;
12 – gyrus temporalis inferior; 13 – parietální lalůček; 14 – postcentrální sulcus; 15 – postcentrální gyrus; 16 – gyrus supramarginální;
17 – úhlový gyrus; 18 – týlní lalok; 19 – mozeček; 20 – horizontální trhlina mozečku; 21 – prodloužená míša



