These are the large hemispheres. Functions of the cerebral hemisphere of the forebrain
Large hemispheres large hemispheres
brain, paired formations united by the corpus callosum into the so-called telencephalon. Surface cerebral hemispheres represented by numerous large or small deep convolutions. There are lobes: frontal, parietal, temporal, insular, occipital. The gray matter of the brain, consisting of nerve cells - neurons, forms the cerebral cortex and subcortical ganglia (nodes). White matter is formed by the processes of neurons that make up the brain's pathways.
LARGE HEMISPHERESLARGE HEMISPHERES of the brain, paired formations united by the corpus callosum (cm. Corpus Callosum) in the so-called telencephalon. The surface of the cerebral hemispheres is represented by numerous large or small deep convolutions. There are lobes: frontal, parietal, temporal, insular, occipital. The gray matter of the brain, consisting of nerve cells - neurons, forms the cerebral cortex and subcortical ganglia (cm. GANGLION)(nodes). White matter is formed by the processes of neurons that make up the brain's pathways.
encyclopedic Dictionary . 2009 .
See what “large hemispheres” are in other dictionaries:
The brain has paired formations, united by the corpus callosum in the so-called. telencephalon. The surface of the cerebral hemispheres is represented by numerous large or small deep convolutions. There are lobes: frontal, parietal, temporal, insular... Big Encyclopedic Dictionary
The brain, paired formations united by the corpus callosum in the so-called. telencephalon. The surface of the B. p. is represented by numerous. b. or m. deep convolutions. There are lobes: frontal, parietal, temporal, insular, occipital. Gray in... ... Natural science. encyclopedic Dictionary
LARGE HEMISPHERES OF THE BRAIN- higher parts of the brain, consisting of the superficial layer of the cerebral cortex and the deep parts of the subcortex; cover the cerebellum and brain stem. B. p. g. m. are divided along the midline into right and left hemisphere, which are in the depths... Psychomotorics: dictionary-reference book
The Arctic Ocean, in contrast to the Southern Ocean, is completely Mediterranean in nature. It has natural boundaries over a considerable distance and only in three places does it directly merge with the waters of the Atlantic and Pacific... ...
The Arctic Ocean, in contrast to the southern one, is completely Mediterranean in nature. It has natural boundaries over a considerable distance and only in three places does it directly merge with the waters of the Atlantic and Pacific... ... Encyclopedic Dictionary F.A. Brockhaus and I.A. Ephron
Also called comparative morphology, this is the study of the patterns of structure and development of organs by comparing various types Living creatures. Data comparative anatomy traditional basis of biological classification. Under morphology... Collier's Encyclopedia
Sectional view of the adult male brain. The human brain (lat. encephalon) is about ... Wikipedia
The science that studies the structure of the body individual organs, tissues and their relationships in the body. All living things are characterized by four characteristics: growth, metabolism, irritability and the ability to reproduce themselves. The combination of these characteristics... ... Collier's Encyclopedia
Animals (Mammalia), the class of vertebrates, the most famous group animals, including more than 4,600 species of the world's fauna. It includes cats, dogs, cows, elephants, mice, whales, people, etc. In the course of evolution, mammals have realized the widest... ... Collier's Encyclopedia
I Medicine Medicine system scientific knowledge and practical activities, the goals of which are to strengthen and preserve health, prolong the lives of people, prevent and treat human diseases. To accomplish these tasks, M. studies the structure and... ... Medical encyclopedia
The cerebral hemispheres are the largest areas of the brain. Human hemispheres big brain received maximum development compared to other parts, which significantly distinguishes the human and animal brains. The left and right hemispheres of the brain are separated from each other by a longitudinal fissure running along the midline. If you look at the surface of the brain from above and from the side, you can see a slit-like depression that begins 1 cm posterior to the midpoint between the anterior and posterior poles of the brain and goes deep. This is the central (Rolandic) sulcus. Below it, along the lateral surface of the brain runs the second large slitlateral (Sylvian) fissure. Functions cerebral hemisphere forebrain- topic of the article.
1 106654
Photo gallery: Functions of the cerebral hemisphere of the forebrain
Lobes of the brain
The large hemispheres are divided into lobes, the names of which are given by the bones covering them: The frontal lobes are located in front of the Rolandic fissure and above the Sylvian fissure.
The parietal lobe lies behind the central lobe and above the posterior portion lateral sulcus; it extends back to the parieto-occipital groove - the gap separating parietal lobe from the occipital, which forms the back of the brain.
The temporal lobe is the area located under the Sylvian fissure and bordered posteriorly by occipital lobe y.
As the brain grows rapidly even before birth, the cerebral cortex begins to increase its surface, forming folds, which leads to the formation of a characteristic appearance brain resembling Walnut. These folds are known as gyri, and the depressions separating them are called fissures. Certain grooves are located in the same place in all people, so they are used as landmarks for dividing the brain into four lobes.
Development of gyri and sulci
Furrows and convolutions begin to appear at the 3-4th month of fetal development. Until this point, the surface of the brain remains smooth, like the brain of birds or amphibians. The formation of a folded structure provides an increase in the surface area of the cerebral cortex in conditions of limited volume of the cranium. Different areas of the cerebral cortex perform specific, highly specialized functions. The cerebral cortex can be divided into the following areas:
Motor areas - initiate and control body movements. The primary motor area controls voluntary movements of the opposite side of the body. Directly in front of the motor cortex is the so-called premotor cortex, and the third area, the supplementary motor area, lies on the inner surface of the frontal lobe.
The sensory areas of the cerebral cortex perceive and synthesize information from sensory receptors throughout the body. The primary somatosensory area receives information from the opposite side of the body in the form of impulses from sensory receptors for touch, pain, temperature, and the position of joints and muscles (proprioceptive receptors).
The surface of the human body has its own “representations” in the sensory and motor areas of the cerebral cortex, which are organized in a certain way. Canadian neurosurgeon Wilder Penfield, who practiced in the 1950s, created a kind of map sensory zones of the cerebral cortex, which perceive information from various parts of the body. As part of his research, he conducted experiments in which he asked a person under local anesthesia to describe his sensations at the moment when he stimulated certain areas of the surface of the brain. Penfield found that stimulation of the postcentral gyrus produced tactile sensations in specific areas on the opposite side of the body. Other studies have shown that the volume of the motor cortex, which is responsible for various areas of the human body, is to a greater extent depends on the level of complexity and accuracy of the movements performed rather than on strength and volume muscle mass. The cerebral cortex consists of two main layers: gray matter - a thin layer of nerve and glial cells about 2-A mm thick and white matter, which is formed by nerve fibers (axons) and glial cells.
The surface of the cerebral hemispheres is covered with a layer of gray matter, the thickness of which in different parts of the brain ranges from 2 to 4 mm. Gray matter is formed by the bodies of nerve cells (neurons) and glial cells that perform a supporting function. In most of the cerebral cortex, under a microscope, six individual layers cells.
Neurons of the cerebral cortex
- Pyramidal cells get their name from the shape of the neuron body, which resembles a pyramid; their axons (nerve fibers) emerge from the cerebral cortex and carry information to other parts of the brain.
- Non-pyramidal cells (all others) are designed to perceive and process information from other sources.
The thickness of the six layers of cells that make up the cerebral cortex varies greatly depending on the region of the brain. German neurologist Korbinian Brodmann (1868-191) examined these differences by staining nerve cells and viewing them under a microscope. The result of Brodmann's scientific research was the division of the cerebral cortex into 50 separate areas based on certain anatomical criteria. Subsequent studies showed that the “Brodmann fields” identified in this way play a specific physiological role and have unique ways of interaction.
Brain (continued)
The greatest size and complexity in mammals is the forebrain, or telencephalon, consisting of two cerebral hemispheres (hemispheri cerebri). Apparently, the hemispheres arose mainly (and perhaps exclusively) in connection with olfactory reception. Smells do not mean much in the lives of higher primates, including humans. However, in earlier stages of evolution, right up to the ancestors of vertebrates, the sense of smell was the main channel, through which animals received information about the world around them. Therefore, it is quite natural that the olfactory centers of the brain served as the basis on which complex nervous mechanisms subsequently developed. Already at the early stages of tetrapod evolution, the hemispheres turn into large and important centers for the correlation of sensory signals. By the time mammals appeared, the greatly expanded surface of the hemispheres became the dominant associative center, the location of higher mental activity. In different representatives of the class, the ratio of the mass of the forebrain hemispheres to the mass of the entire brain varies: in the hedgehog ( Erinaceus europaeus) it is 48%, for squirrels ( Sciurus vulgaris) - 53%, in the wolf ( Canis lupus) - 70%, in the white sided ( Delphinus delphis) - 75%, in most primates - 75-80%, in humans - about 85%. In birds, the cerebral hemispheres are approximately equal in mass to the rest of the brain or inferior to it, sometimes several times. Finally, the extreme importance of the cerebral hemispheres is evidenced by the fact that their destruction leads to complete functional failure of the mammal.
The olfactory bulbs (bulbi olfactorii) are adjacent to the anterior part of the hemispheres. These formations are most developed in animals with a good sense of smell and are greatly reduced in purely aquatic forms. Based on the differences in their development, several types of brain structure are distinguished. In marsupials, insectivores, edentates, carnivores, rodents and some others, the olfactory bulbs are large and protrude well when looking at the brain from above. This type of brain with perfect development of the olfactory lobes is called macrosmatic. In pinnipeds, sirenians and many primates, the bulbs are poorly developed; These animals are characterized by a microsmatic brain. Finally, for cetaceans the so-called anosmatic brain with reduced olfactory bulbs. It was previously thought that the ability to distinguish between chemical signals was completely lost in whales and dolphins, but it turns out that this is not entirely true.
The superficial layers of the mammalian forebrain hemispheres form the pallium, or the medullary vault (pallium). The upper layer, consisting of neuron bodies and soft nerve fibers, is called the cortex (cortex cerebri) and represents the gray matter of the fornix. The bodies of neurons are located in the cortex in layers, forming peculiar screen structures. This organization of the brain allows the external world to be spatially represented based on information coming from the senses. Screen structures are characteristic of the most important brain centers of mammals, while in other vertebrates they are less common, mainly in the visual centers. Under the cortex there is a layer of myelinated neuron processes - the white matter of the fornix. The nerve fibers of the white matter form conductive bundles that solder the hemispheres - cerebral commissures. Although the main part of the gray matter is concentrated in the cortex, it is also present in the subcortical layer, where it is represented by relatively small clusters - nuclei. These include the striatal bodies (corpora striata), which lie under the cavity of the lateral ventricle and received their name because of the nerve fibers crossing them. Under the control of the cortex, the striatum performs the function of regulating stereotypical, automated reactions - unconditioned reflexes.
Schemes reflecting the progressive development of the forebrain hemispheres. Side view of the hemisphere with the olfactory bulb. Different sections that differ from each other cytologically are stained differently.
A - the hemisphere is just the olfactory lobe.
B - the dorsal section - the archipallium (= hippocampus) and the ventral section - the basal nucleus (striatum) are differentiated.
B - the basal ganglia moved to the inner region of the hemisphere.
D - a small area appears, representing a neopallium.
D - the paleopallium is pushed onto the medial surface of the hemisphere, but the neopallium is still modest in size, and significantly developed olfactory sections are preserved under the olfactory sulcus.
E - the primitive olfactory department is preserved only in the ventral region, and the neopallium reaches an extremely strong development. (After Romer and Parsons, 1992.) The brain vault began to form in fish. Due to the progressive development of the olfactory sensory system, they develop a paleopallium, or ancient arch (paleopallium), completely covering the small hemispheres. At the stage of lobe-finned fish, the archipallium, or old arch (archipallium), appears in the dorsal part of the hemispheres closer to the axis of the body. In amphibians and primitive reptiles it receives further development, as a result of which the paleopallium is pushed aside and is preserved only along the lateral surface of the hemispheres. At the same time, the paleopallium continues to be predominantly olfactory in nature and at higher stages of evolution forms the olfactory lobes of the cerebral cortex. The archipallium is to a certain, albeit small, extent a correlative center, receiving ascending fibers from the diencephalon, as well as fibers from the olfactory bulb and olfactory lobe; it also appears to be associated with emotional behavior. The neural pathway from this region to the hypothalamus forms the main element of a bundle of fibers called the fornix in mammals.
In amphibians, the rudiment of the neopallium, or new arch (neopallium), appears for the first time. In reptiles, the neopallium already forms a small area between the ancient and old vault. From the very beginning of its evolution, this area has been an associative center, like the basal ganglia, receiving fibers that switch sensory signals from the brain stem to it and, conversely, transmit commands directly to the motor columns.
In monotremes, the neopallium is still wedged between the paleopallium on the outside and the archipallium on the inside. In marsupials, it grows beyond the roof and side walls of the hemispheres. In this case, the archipallium is displaced onto the medial surface, and the paleopallium is limited to the ventrolateral part of the hemisphere, located below the nasal groove (fissura rhinalis) - a groove that represents the boundary between the olfactory and non-olfactory areas of the cortex. In placentals, due to further complication and expansion of the neopallium, the hemispheres reach such a size that they exceed in volume the rest of the brain parts combined. The hemispheres grow back and to the sides, successively covering the diencephalon, midbrain and part of the cerebellum. The paired ventricles and ancient structures primarily responsible for the sense of smell (olfactory bulbs, old layers of the medullary vault and associated nerve bundles and nuclei) are pushed back and deformed in the placental brain. Thus, the paleopallium is preserved on the ventral surface of the hemisphere in the form of a small olfactory area called the piriform lobe (lobus piriformis), and the archipallium is folded deep into the fold of the temporal lobe into a roll called the hippocampus (hippocampus). The superiority of the cerebral hemispheres over other parts of the brain is noticeable in all mammals, but it is especially pronounced in advanced forms, such as humans. The hemispheres also dominate functionally. Unlike mammals, the complication of the forebrain in birds is expressed mainly in the growth of the basal nuclei (nuclei basales), rather than in the remaining thin fornix. 
Schematic sections through the left forebrain hemisphere. The color designations are the same as in the previous figure.
1 - paleopallium; 2 - lateral ventricle; 3 - archipallium; 4 - basal nuclei; 5 - neopallium; 6 - corpus callosum.
A - primitive stage. The hemisphere, in essence, is the olfactory lobe. Poorly differentiated gray matter is located inside the brain.
B - stage observed in modern amphibians. The gray matter is still located far from the outer surface, but is already divided into paleopallium (= olfactory lobe), archipallium (= hippocampus) and basal ganglia (= striatum). The latter acquires the significance of an associative center that has afferent and efferent connections with the thalamus (depicted by lines symbolizing cut bundles of fibers).
B - a more progressive stage, at which the basal ganglia are immersed inside the hemisphere, while the sections of the cortex have moved somewhat outward.
G is the stage at which advanced reptiles are located. Neopallium appears.
D - stage of a primitive mammal. Neopallium increased. It has extensive connections with the brain stem. Archipallium on medial surface hemisphere wraps up as the hippocampus. The paleopallium is still highly developed.
E - stage of a highly organized mammal. Neopallium grows extremely and gathers in folds. The paleopallium occupies a limited ventral region, representing a pear-shaped lobe. The corpus callosum develops - a powerful bridge connecting the neopallium areas of the two hemispheres. (After Romer and Parsons, 1992.) The cortex of the neopallium is called the neocortex, or neocortex. In mammals, it serves as the center of the higher (conditioned reflex) nervous activity, coordinating the work of other parts of the brain. From here impulses are sent to various organs and tissues of the body, and here the regulation of physiological processes is carried out in accordance with environmental conditions. It is the new cortex that accumulates traces of single excitations and their combinations, as a result of which the operative memory is enriched, providing the ability to choose optimal solutions in new situations. More often than not, these decisions represent new combinations of previously known behavioral elements, but new options for action are also developed and consolidated. As it develops, the new cortex not only takes on the functions of a correlative and associative center of newly emerging types of higher nervous activity, but also begins to perform many functions that previously belonged to the centers of the brain stem and basal ganglia. In this case, the ancient centers that control instinctive acts are not eliminated, but are only subject to higher control.
In connection with the development of the new cortex, the roof of the midbrain loses its former significance, remaining only a reflex and transmission center. Auditory and other somatic sensory impulses are transmitted forward to the thalamus, where most of the visual fibers are interrupted, and all these signals from the thalamus are transmitted to the hemispheres along powerful nerve bundles. Similar thalamic connections with the basal ganglia arose in low-organized groups of vertebrates and were most developed in birds. Unlike birds, in mammals the bulk of the fibers pass through the striatum and diverge to the surface of the neocortex. Thus, a complete set of sensory data flows to it, on the basis of which the corresponding motor “decisions” are made in the cortex.
As already mentioned, some signals are transmitted from the cortex to the cerebellum through the pons and provide the necessary regulatory effects. The neocortex also has connections with the striatum and even with the hypothalamus - and thus with the autonomic nervous system. However, the bulk of motor commands are sent along the pyramidal tract (tractus corticospinalis) - a special nerve bundle that directly, without switching, goes from the cerebral cortex through the midbrain to the somatic motor areas of the brain stem and spinal cord. In this case, the fibers of the lateral part of this path intersect and innervate the opposite side of the body (i.e., the left fiber innervates the right part of the body, and vice versa), and the ventral fibers remain connected to their side of the body. The pyramidal tract is present only in mammals, which clearly demonstrates the dominant position of the neocortex in them. Greatest development this structure reaches in monkeys and, especially, in humans, by playing important role in upright posture. In marsupials, pyramidal axons reach only the thoracic region, while in monotremes, the pyramidal tract is completely absent. 
Ventricles of the human brain; left side view. The ventricles are represented as a cast, but the brain tissue is not depicted. With the growth of the forebrain hemisphere, the lateral ventricle spread backward to form the posterior horn in the occipital lobe, and in its lateral part - down and forward to form the lateral horn in the temporal lobe. These growths, directed backwards and downwards, led to changes in the arrangement of various parts of the brain. The hippocampus, which developed in a dorsal position on the medial surface of the hemisphere, moved back and down to a ventral position in highly developed mammals. (After Romer and Parsons, 1992.) Since the neocortex is a thin sheet of layered cellular material underlying the white fibrous mass of the brain, simply increasing the volume of the hemispheres cannot produce proportional growth of the cortex. At the same time, in advanced forms the area of the cortex can increase significantly due to its folding. The folds formed in this way are called convolutions (gyri), and the deep gaps between them are called grooves (sulci). Both contain common morphological components. In the simplest case, there is one deep Sylvian fissure separating the frontal lobe (lobus frontalis) from the temporal lobe (lobus temporalis). Then, above and anterior to the Sylvian fissure, a transversely running Rolandic fissure appears, separating the frontal lobe from the parietal lobe (lobus parietalis) above. In primates, a transverse groove separates the small posterior occipital lobe (lobus occipitalis). In addition to the main furrows, many additional furrows are formed; their numbers are especially high in primates and toothed whales. Previously, it was believed that grooves in some cases mark morphological boundaries corresponding to certain areas of the cortex. However, further studies showed the absence of a fixed connection between the distribution of folding and structural unit cortex (except for the nasal sulcus and, to some extent, the central sulcus in primates, which will be discussed later). It is noteworthy that cortical folding developed completely independently in several evolutionary trunks of mammals. In relatively primitive mammals, such as monotremes, marsupials, as well as some placentals (insectivores, chiropterans, rodents, lagomorphs), the cortex is more modestly developed and has a smooth surface. 
The location of the brain in the skull of fossil and living canids. There is a noticeable increase in the size and complexity of the brain, especially the forebrain hemispheres. Hesperocyon ( Hesperocyon gregarius) (left) - an Oligocene form that lived approximately 30 million years ago. Fennec ( Vulpes zerda) (on right) - modern form similar sizes. (After Romer and Parsons, 1992.) The gray matter of the neocortex is characterized by a complex histological structure. In placental mammals, there are 6 layers of cells lying on top of each other and fibers embedded between them; this greatly distinguishes the neocortex from the preserved areas of the cortex of the paleopallium and archipallium, where only 2 to 4 layers of cells can be distinguished. Current estimates suggest that in mammals, which have particularly large brains, the number of neocortical cells may reach billions.
The white matter, located under the gray matter, in addition to the fan of connections going from the cortex to the underlying parts of the brain and back, includes great amount interwoven transverse fibers connecting different areas of the cortex itself. The commissure thus formed is stretched backwards (according to the growth rate of the hemispheres) and is divided into two plates fused along the posterior edge. The lower one, thinner and deflected downwards by the anterior edge, is the fornix (fornix), the commissure of the archipallium cortex (i.e., the hippocampus). The upper, thicker horizontal commissure belongs to the new cortex and is called the corpus callosum. This education allows you to combine the memory of both hemispheres and significantly increases the brain's ability to learn. The corpus callosum is present only in placentals due to the significant development of the neocortex; monotremes and marsupials lack it. In addition, all mammals have an anterior commissure (commissura anterior), connecting the olfactory areas of the cortex. 
Layer-by-layer arrangement of nerve cells in the cortex telencephalon mammal (according to Naumov and Kartashev, 1979.) A complex system“conductors” connecting all parts of the cortex suggests that the gray matter is, in principle, a single formation, all parts of which have the same capabilities to carry out any functions of the cerebral hemispheres. To a certain extent, this is true: experiments show that in laboratory animals it is possible to destroy a significant part of the neocortex without causing permanent disturbances in their normal activity. Data on injury and morbidity confirm that this is also true for human brain. At the same time, it is clear that certain areas of the cortex are normally associated with the performance of very specific functions. Mentioned above were the areas of the paleopallium and archipallium, intended mainly for the analysis of olfactory information and preserved, respectively, in the form of the piriform lobe and hippocampus. Differentiation of individual areas also takes place in the cortex of the neopallium. The anterior part of the hemispheres contains the motor area. The frontal lobe located here, among other things, controls animal communication, including acoustic communication; in humans it is associated with speech, i.e. the second alarm system. The back of the hemispheres is associated with the perception of sensations. The occipital and temporal lobes contain areas that control vision and hearing, respectively. Further forward, near the motor area, there are areas that receive tactile and proprioceptive signals. In primates, the central sulcus (sulcus centralis), crossing the apex of the hemisphere from the medial to the lateral surface, delimits (although not entirely accurately) the motor area from the sensory area. Along the anterior edge of the central sulcus, specific motor areas serving each part of the body and limbs are located in a linear order. Along the posterior edge of the central sulcus, areas of sensory perception of the corresponding parts of the body are located in the same order.
Thus, in many mammals, almost the entire surface of the neocortex is occupied by areas more or less closely associated with certain sensory or motor functions. Although the central sulcus may be absent, placentals generally maintain a similar linear arrangement of sensory and motor areas opposite each other. In marsupials (and among placentals, in xenarthrids), the “marking” of body areas is approximately the same, but the sensory areas are not separated from the motor ones, but alternate with them. But, for example, in humans, these specific functional areas occupy relatively little space on the surface of the neocortex. Between them, large areas of gray matter have arisen (one particularly large such area occupies most of the frontal lobe) that are not associated with specific sensory or motor functions. Therefore, these areas are often called “white spots,” although, as damage to these areas shows, they are where our higher mental abilities are located, including learning, initiative, foresight, and judgment. At the same time, there are also areas that can be removed without serious consequences for intellectual activity. 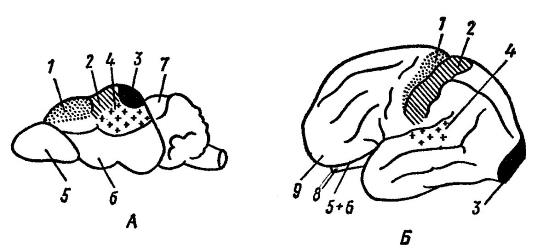
Functional centers of the shrew's cerebral cortex ( Sorex sp.) (A) and human ( Homo sapiens) (B) (according to Naumov and Kartashev, 1979):
1 - motor center; 2 - center of musculocutaneous sensitivity; 3 - visual center; 4 - auditory center; 5 - olfactory bulb; 6 - olfactory lobes; 7 - roof of the midbrain; 8 - cerebellum; 9 - frontal lobe. On the evolution of the brain big influence exerted by the external environment and motor (food-procuring, defensive) activity. At the same time, the development of various parts of the brain is determined mainly by the methods of finding food: in a dog ( Canis lupus), using the sense of smell in this process, the olfactory area is more developed; in a cat ( Felis silvestris), searching for food using vision - visual; in the macaque ( Macaca mulatta), using vision and hearing - visual and auditory.
It is usually assumed that the size of the cerebral hemispheres determines differences in the mental abilities of different mammals. IN in a certain sense this is true, but with significant reservations. The larger brain is made up of more nerve cells. If available cortical surface area is in any way related to intelligence, then it is clear that of the two brain variants same sizes the more developed will be the one with a grooved surface, and the less developed will be the brain with a smooth surface. The size of the animal itself also affects the volume of the brain. This happens if only because the brain must have larger areas to serve more extensive sensory and motor connections. However, the increase in brain size is not entirely proportional to body mass, so that large animals tend to have relatively smaller brains without any apparent loss of mental capacity. Thus, the absolute size of the brain is not an absolute criterion of intelligence. This is certainly indicated by the fact that the whale brain can be five times larger in volume than the human brain.
Comparison of the brains of some mammals:
1 - horse; 2 - dog; 3 - kangaroo; 4 - person; 5 - elephant. The percentage of the brain in the total body mass is called the cephalization index. In large insectivores it is about 0.6%, in small ones - up to 1.2%, in large cetaceans - about 0.3%, and in small ones - up to 1.7%. In most primates, the cephalization index is 1-2%. In humans it reaches 2-3%, and some small broad-nosed monkeys have a brain whose mass makes up up to 7% of their body weight. At the same time, in modern reptiles and birds the cephalization index ranges from 0.05 to 0.5%.
The following is the brain mass of some mammals (the mass of the animal is indicated in parentheses):
Virginia opossum ( Didelphis virginiana) - 7.6 g (5 kg);
koala ( Phascolarctos cinereus) - 19.2 g (8 kg);
savannah elephant ( Loxodonta africana) - 6000 g (5000 kg);
common hedgehog ( Erinaceus europaeus) - 3.3 g (1 kg);
house mouse ( Mus musculus) - 0.3 g (0.02 kg);
gray rat ( Rattus norvegicus) - 2 g (0.3 kg);
common squirrel ( Sciurus vulgaris) - 7 g (0.4 kg);
European rabbit ( Oryctolagus cuniculus) - 11 g (3 kg);
domestic horse ( Equus ferus) - 530 g (500 kg);
black rhino ( Diceros bicornis) - 500 g (1200 kg);
white-tailed deer ( Odocoileus virginianus) - 500 g (200 kg);
giraffe ( Giraffa camelopardalis) - 680 g (800 kg);
domestic sheep ( Ovis orientalis) - 140 g (55 kg);
domestic bull ( Bos primigenius) - 490 g (700 kg);
Bactrian camel ( Camelus bactrianus) - 762 g (700 kg);
hippopotamus ( Hippopotamus amphibius) - 580 g (3500 kg);
common dolphin ( Delphinus delphis) - 815 g (60 kg);
narwhal ( Monodon monoceros) - 2997 g (1578 kg);
sperm whale ( Physeter macrocephalus) - 8028 g (35833 kg);
blue whale ( Balaenoptera musculus) - 3636 g (50900 kg);
domestic cat ( Felis silvestris) - 25 g (3 kg);
a lion ( Panthera leo) - 270 g (250 kg);
common fox ( Vulpes vulpes) - 53 g (4.5 kg);
domestic dog ( Canis lupus) - 64 g (10 kg);
polar bear ( Ursus maritimus) - 500 g (700 kg);
walrus ( Odobenus rosmarus) - 1130 g (700 kg);
marmoset Geldi ( Callimico goeldii) - 7 g (0.2 kg);
white-fronted capuchin ( Cebus albifrons) - 57 g (1 kg);
rhesus macaque ( Macaca mulatta) - 88 g (6.5 kg);
baboon ( Papio cynocephalus) - 200 g (25 kg);
silver gibbon ( Hylobates moloch) - 112 g (6.5 kg);
Kalimantan orangutan ( Pongo pygmaeus) - 413 g (50 kg);
western gorilla ( Gorilla gorilla) - 506 g (126 kg);
common chimpanzee ( Pan troglodytes) - 430 g (55 kg);
reasonable person ( Homo sapiens) - 1400 g (72 kg).
From the above examples it is clear that in smaller mammals the brain is almost always relatively larger, and as the size of the animal’s body increases, the relative size of the brain decreases. This is especially pronounced among closely related species of mammals - for example, in cats ( Felis silvestris) and lion ( Panthera leo). Dogs are also very convincing in this sense. various breeds. If the body masses of the smallest and largest breeds are approximately in the ratio of 1:33, then the brain masses of the same breeds are in the ratio of 1:3. 
Ranges of brain and body mass values for some groups of vertebrates. In domesticated animals, deprived of the need to obtain food and defend themselves from enemies, the size of the brain is significantly reduced. For example, the volume of a wolf's brain ( Canis lupus) 30% more than a dog of the same size. Interestingly, these changes affect not only traditionally domestic animals, but also representatives of free-living species kept in captivity for some time. Yes, foxes ( Vulpes vulpes), born in nature, but from the very first days living in captivity, have a smaller brain than their relatives living in natural conditions. In this case, the differences reach 20%, which approximately corresponds to the difference in brain volume of wild and real domestic animals. Brain shrinkage, although not as pronounced (at approximately 5%), has been found in captive wolves ( Canis), ferrets ( Mustela), rats ( Rattus). In this case, the decrease does not affect all parts of the brain, but only those areas that are associated with the functioning of the sensory organs. The most remarkable thing is that the brain weight of released domestic animals increases. For example, feral cats have brains that are about 10% larger than their domesticated counterparts. Significant brain enlargement was also found in feral rabbits ( Oryctolagus cuniculus) on the Kerguelen Islands. Feral donkeys ( Equus asinus) V South America had 15% big brain than homemade ones. It is also interesting that the brains of a Neanderthal ( Homo neanderthalensis) and Paleolithic homo sapiens ( Homo sapiens) were somewhat larger in size than the brain of modern humans.
It has been established that many mammals exhibit motor asymmetry, that is, the predominant use of the right or left half of the body. For example, when studying unbroken horses ( Equus ferus) recorded which leg the animals start walking with, which side they prefer to go around obstacles, and which side they prefer to lie on in a stall on hay. As a result, most mares were right-handed, and most stallions were left-handed. Approximately 10% of horses showed no preference for either the right or left limb. According to observations, about 90% of walruses ( Odobenus rosmarus) dig shellfish out of sea mud with the right flipper. Cradling babies, about 80% of female chimpanzees ( Pan) and gorillas ( Gorilla) press their heads to the left side of their chest (about the same percentage is observed among women). Rats ( Rattus), searching for food using vibrissae located on right side muzzles, are more prey than their left-handed relatives.
text_fields
text_fields
arrow_upward
The diencephalon, together with the brain stem, is covered from above and from the sides large hemispheres - telencephalon. The hemispheres consist of subcortical ganglia (basal ganglia), and have cavities -. The outside of the hemispheres are covered (with a cloak).
Basal ganglia or subcortical ganglia
text_fields
text_fields
arrow_upward
Basal ganglia or subcortical nodes (nuclei basales)– formations are phylogenetically more ancient than the bark. The basal ganglia got their name due to the fact that they lie at the base of the cerebral hemispheres, in their basal part. These include the caudate and lenticular nuclei, united in the striatum, the fence and the amygdala.
Caudate nucleus
text_fields
text_fields
arrow_upward
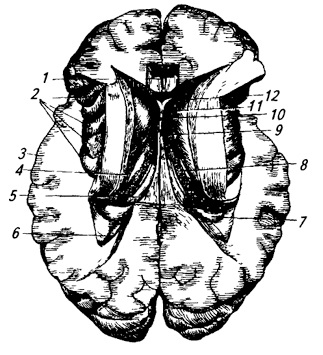
Caudate nucleus (nucleus caudatus) elongated in the sagittal plane and strongly curved (Fig. 3.22; 3.32; 3.33). Its anterior, thickened part is head– is located in front of the optic thalamus, in the lateral wall of the anterior horn of the lateral ventricle, behind it gradually narrows and turns into tail. The caudate nucleus covers the visual thalamus in front, above and on the sides.
Rice. 3.22.
1 – caudate nucleus;
2 – columns of the vault;
3 – pineal gland;
4 – upper and
5 – inferior colliculus;
6 – fibers of the middle cerebellar peduncle;
7 – pathway of the superior cerebellar peduncle (prepared);
8 – tent core;
9 – worm;
10 – spherical,
11 – corky and
13 – dentate nucleus;
12 – cerebellar cortex;
14 – superior cerebellar peduncle;
15 – leash triangle;
16 – thalamic cushion;
17 – visual thalamus;
18 – posterior commissure;
19 – third ventricle;
20 – anterior nucleus of the visual thalamus
 Rice. 3.32.
Rice. 3.32. Rice. 3.32. Brain - horizontal section through the lateral ventricles:
1 – corpus callosum;
2 – island;
3 – bark;
4 – tail of the caudate nucleus;
5 – vault;
6 – posterior horn of the lateral ventricle;
7 – hippocampus;
8 – choroid plexus;
9 – interventricular foramen;
10 – transparent partition;
11 – head of the caudate nucleus;
12 – anterior horn of the lateral ventricle
Lentil shaped core
text_fields
text_fields
arrow_upward
Lentil shaped core (nucleus lentiformis) located outside the optic thalamus, at the level of the insula. The shape of the core is close to a triangular pyramid, with its base facing outward. The nucleus is clearly divided by layers of white matter into a darker-colored lateral part - shell and medial - pale ball, consisting of two segments: internal and external (Fig. 3.33; 3.34).
 Rice. 3.33.
Rice. 3.33. Rice. 3.33. Horizontal section of the cerebral hemispheres at the level of the basal ganglia:
1 - corpus callosum;
2 – vault;
3 – anterior horn of the lateral ventricle;
4 – head of the caudate nucleus;
5 – internal capsule;
6 – shell;
7 – globus pallidus;
8 – outer capsule;
9 – fence;
10 – thalamus;
11 – pineal gland;
12 – tail of the caudate nucleus;
13 – choroid plexus of the lateral ventricle;
14 – posterior horn of the lateral ventricle;
15 – cerebellar vermis;
16 – quadrigeminal;
17 – posterior commissure;
18 – cavity of the third ventricle;
19 – pit of the lateral groove;
20 – island;
21 – anterior commissure
 Rice. 3.34.
Rice. 3.34. Rice. 3.34. Frontal section through the cerebral hemispheres at the level of the basal ganglia:
1 - corpus callosum;
2 – lateral ventricle;
3 – caudate nucleus (head);
4 – internal capsule;
5 - lenticular shaped core;
6 – lateral groove;
7 - temporal lobe;
8 – fence;
9 – island;
10 – outer capsule;
11 – transparent partition;
12 – radiance of the corpus callosum;
13 – cerebral cortex
Shell
text_fields
text_fields
arrow_upward
 Rice. 3.35.
Rice. 3.35. Shell (putamen) according to genetic, structural and functional characteristics it is close to the caudate nucleus.
Both of these formations have a more complex structure than the globus pallidus. Fibers approach them mainly from the cerebral cortex and thalamus (Fig. 3.35).
Rice. 3.35. Afferent and efferent connections of the basal ganglia:
1 - precentral gyrus;
2 – shell;
3 – outer and inner segments of the globus pallidus;
4 – lenticular loop;
5 - reticular formation;
6 – reticulospinal tract,
7 - rubrospinal tract;
8 – cerebellothalamic tract (from the dentate nucleus of the cerebellum);
9 – red core;
10 – substantia nigra;
11 – subthalamic nucleus;
12 – Zona incerta;
13 – hypothalamus;
14 – ventrolateral,
15 – intralaminar and centromedian nuclei of the thalamus;
16 – III ventricle;
17 – caudate nucleus
Pale ball
text_fields
text_fields
arrow_upward
The globus pallidus (globus pallidus) is mainly associated with the conduction of impulses along numerous descending pathways into the underlying structures of the brain - the red nucleus, substantia nigra, etc. Fibers from the neurons of the globus pallidus go to the same nuclei of the thalamus that are connected to the cerebellum. From these nuclei, numerous pathways go to the cerebral cortex.
The globus pallidus receives impulses from the caudate nucleus and putamen.
The striatum (corpus striatum), which unites the caudate and lentiform nuclei, belongs to the efferent extrapyramidal system. The dendrites of striatal neurons are covered with numerous spines. Fibers from neurons of the cortex, thalamus and substantia nigra terminate on them (Fig. 3.35). In turn, striatal neurons send axons to the intralaminar, anterior and lateral nuclei of the thalamus. From them, the fibers go to the cortex, and thus the feedback loop between the cortical neurons and the striatum is closed.
During the process of phylogenesis, these nuclei were built on top of the nuclei of the midbrain. Receiving impulses from the thalamus, the striatum takes part in such complex automatic movements as walking, climbing, and running. In the nuclei of the striatum, the arcs of the most complex unconditional ones are closed, i.e. congenital reflexes. The extrapyramidal system is phylogenetically more ancient than the pyramidal system. In a newborn, the latter is not yet sufficiently developed and impulses to the muscles are delivered from the subcortical ganglia through the extrapyramidal system. As a result, the child’s movements in the first months of life are characterized by generalization and undifferentiation. As the cerebral cortex develops, the axons of their cells grow to the basal ganglia, and the activity of the latter begins to be regulated by the cortex. The subcortical ganglia are associated not only with motor reactions, but also with autonomic functions - these are the highest subcortical centers autonomic nervous system.
Amygdala
text_fields
text_fields
arrow_upward
Amygdala (corpus atugdaloideum) (amygdala) – a collection of cells in the white matter of the temporal lobe. With help anterior commissure it connects with the body of the same name on the other side. The amygdala receives impulses from a variety of afferent systems, including the olfactory system, and is related to emotional reactions(Fig. 3.36).
 Rice. 3.36.
Rice. 3.36. Rice. 3.36. Brain structures associated with the amygdala: afferent (A) and efferent (B) connections of the amygdala:
1 - thalamic nuclei;
2 – periaqueductal gray matter;
3 – parabrachial nucleus;
4 – blue spot;
5 - suture cores;
6 – nucleus of the solitary tract;
7 - dosal nucleus of the X nerve;
8 – temporal cortex;
9 – olfactory cortex;
10 – olfactory bulb;
11 - frontal cortex;
12 – cingulate gyrus;
13 – corpus callosum;
14 – olfactory nucleus;
15 - anteroventral and
16 – dorsomedial nucleus of the thalamus;
17 – central,
18 – cortical and
19 – basolateral nucleus of the amygdala;
20 – hypothalamus;
21 – reticular formation;
22 – partition;
23 – substantia nigra;
24 – ventromedial nucleus of the hypothalamus; XXIII, XXIV, XXVIII – cortical fields
>>Forebrain functions
§ 46. Functions of the forebrain
1. Where are perceived images recognized?
2. Do the left and right hemispheres perform the same functions?
The forebrain consists of two sections: the diencephalon and the cerebral hemispheres brain. This is the largest section of the brain, consisting of the right and left halves.
The diencephalon consists of three parts - upper, central and lower (Fig. 93-97). The central part of the diencephalon is called the thalamus. It consists of two paired formations separated by the third ventricle of the brain. Everyone flocks here information from the senses. Here comes the first assessment of its significance. Thanks to the thalamus only important information enters the cerebral cortex.
The lower part of the diencephalon is called the hypothalamus. It regulates metabolism and energy. In its cores there are centers of thirst and its quenching, hunger and saturation. The hypothalamus controls the satisfaction of needs and maintaining a constant internal environment - homeostasis. With the participation of the diencephalon and other parts of the brain, many cyclic movements are carried out: walking, running, jumping, swimming, etc., as well as maintaining posture between movements.
The cerebral hemispheres are divided by a deep anteroposterior fissure into left and right parts. In its depths there is a bridge of white matter connecting them - the corpus callosum.
The surface of the cerebrum is formed by the cortex, consisting of gray matter. The bodies of neurons are concentrated there. They are arranged in columns, forming several layers.
The cortex is a white substance consisting of a mass nerve fibers, connecting cortical neurons with each other and with underlying parts of the brain. In the thickness of the hemispheres, among the white matter, there are islands of gray matter in the form of nuclei, forming subcortical centers.
The surface of the hemispheres is folded. The protruding parts of the surface form convolutions, and the depressions form grooves. They greatly increase the surface area of the cerebral cortex. The deepest grooves divide each hemisphere into four lobes: frontal, parietal, occipital and temporal (Fig. 95). They are adjacent to the corresponding bones and therefore bear their names. The central sulcus separates the frontal lobe from the parietal lobe, the lateral sulcus separates the temporal lobe from the frontal and parietal lobes.
Analysis occurs in the neurons of the cerebral cortex nerve impulses, coming from the senses (Fig. 96). It is carried out in sensitive areas that occupy the middle and back parts of the brain. Thus, neurons are concentrated in the occipital lobe visual area, in the temporal - auditory. In the parietal zone, behind the central gyrus, there is an area of musculocutaneous sensitivity.
The olfactory and gustatory zones are located on the inner surface of the temporal lobes. The centers that regulate active behavior are located in the front parts of the brain, in the frontal lobes of the cerebral cortex.
The motor zone is located in front of the central gyrus.
Right hemisphere controls the organs of the left side of the body and receives information from the space on the left. The left hemisphere regulates the functioning of the organs of the right side of the body and perceives information from space on the right.

The main feature of the large human brain is that the right and left hemispheres are functionally different. In the left hemisphere, as a rule, right-handed people have speech centers. Here the situation and related actions are analyzed according to individual parameters, generalizations are developed, and logical conclusions are drawn. The right hemisphere perceives the situation as a whole. This is where so-called intuitive solutions arise. The right hemisphere recognizes images and melodies and remembers faces.
In the cerebral hemispheres, temporary connections are formed between signal, conditioned reflex stimuli and vital events. Through these connections, individual experience is accumulated.
Old and new cerebral cortex. Old bark is already present in reptiles. In mammals, its appearance is associated with the development of the sense of smell. It surrounds the base of the brain like a belt and includes the subcortical nuclei (Fig. 97).
Centers associated with complex instincts, emotions, and memory are concentrated here. The old cortex allows the body to distinguish between favorable and unfavorable events and respond to them with fear, joy, aggression, and anxiety. This is where information about experienced events is stored in memory. This makes it possible, under similar circumstances, to take actions that will lead to success. Unlike the neocortex, the old cortex cannot accurately recognize objects, estimate the likelihood of future events, and plan responses to their occurrence.
And the new cortex receives information from internal organs and from sensory organs. In the frontal lobes, the most important one is selected from numerous needs and the goal of the activity is formed, a plan for achieving the goal based on an analysis of the situation and past experience.
Here, with the participation of speech centers, scenarios of future behavior are developed. They are implemented by other parts of the brain and spinal cord associated with the executive organs.

Information about the results achieved comes via feedback V frontal lobes hemispheres and, depending on the effect obtained, the activity stops or continues in a modified form.
Forebrain; diencephalon: thalamus, hypothalamus; cerebral hemispheres, corpus callosum, cortex, sulci, gyri, lobes of the brain: sensory and motor areas; temporary (conditioned reflex) connections; old and new bark.
1. What divisions are distinguished in the forebrain?
2. What are the functions of the thalamus and hypothalamus?
3. Why is the surface of the hemispheres folded?
4. How is gray and white matter distributed in the cerebral hemispheres? What functions do they perform?
5. What are the functions of the old cortex?
6. How are functions distributed between the left and right hemispheres of the cerebrum?
7. Which connections in the body are called direct and which are called reverse?
Explain in which hemisphere the damage to the motor centers occurred if the patient experienced paralysis of the right leg or arm.
Kolosov D.V. Mash R.D., Belyaev I.N. Biology 8th grade
Submitted by readers from the website